Анализ кала на дисбактериоз
Анализ кала на дисбактериоз
Представлены обобщенные сведения о традиционном микробиологическом анализе кала культурально-зависимым способом (бакпосевом).
О новых методиках анализа кишечной микробиоты информация ниже.
Анализ кала на дисбактериоз или Бактериологический посев (посев или бакпосев). Посев — это биологическое исследование испражнений, которое определяет состав и примерное количество микроорганизмов, обитающих в кишечнике человека. Для этого используется внесение частиц кала на разные питательные среды, на которых растут 3 группы микроорганизмов: нормальные (необходимы для переваривания пищи), условно-патогенные (способные при снижении естественной резистентности макроорганизма вызывать заболевания, для которых характерно отсутствие нозологической специфичности) и патогенные (болезнетворные).
Данное исследование позволяет определить содержание бактерий в толстом кишечнике. В кишечнике человека содержится огромное количество бактерий, которые активно участвуют в переваривании и усвоении питательных веществ. Анализ кала на дисбактериоз назначается по большей части детям, в таких случаях, когда имеются следующие нарушения работы кишечника: диарея, запор, боли в животе, метеоризм, а также после длительного лечения антибиотиками (антибиотики помимо того, что борются с инфекциями, уничтожают и нормальные бактерии кишечника). Существую три группы бактерий кишечника – «нормальные» бактерии (бифидобактерии, лактобактерии и эшерихии) они активно участвуют в работе кишечника, условно-патогенные бактерии (энтерококки, стафилококки, клостридии, кандиды) при определенных обстоятельствах могут превратиться в патогенные бактерии и вызвать различные заболевания, и патогенные бактерии (шигелла, сальмонелла) которые при попадании в кишечник вызывают серьезные инфекционные заболевания кишечника.
(КОЕ / г фекалий)
| Дети до 1 года | Дети старшего возраста | Взрослые |
Бифидобактерии | 1010 – 1011 | 109 – 1010 | 108 – 1010 |
Лактобактерии | 106 – 107 | 107 – 108 | 106 – 108 |
106 – 107 | 107 – 108 | 106 – 108 | |
Бактероиды | 107 – 108 | 107 – 108 | 107 – 108 |
Пептострептококки | 103 – 105 | 105 – 106 | 105 – 106 |
Энтерококки | 105 – 107 | 105 – 108 | 10 |
Сапрофитные стафилококки | ≤104 | ≤104 | ≤104 |
Патогенные стафилококки | — | — | — |
Клостридии | ≤103 | ≤105 | ≤105 |
Кандида | ≤103 | ≤104 | ≤104 |
Патогенные энтеробактерии | — | — | — |
Бифидобактерии
Норма бифидобактерий
Дети до 1 года | 1010 – 1011 |
Дети старшего возраста | 109 – 1010 |
Взрослые | 108 – 1010 |
Около 95% всех бактерий в кишечнике – бифидобактерии.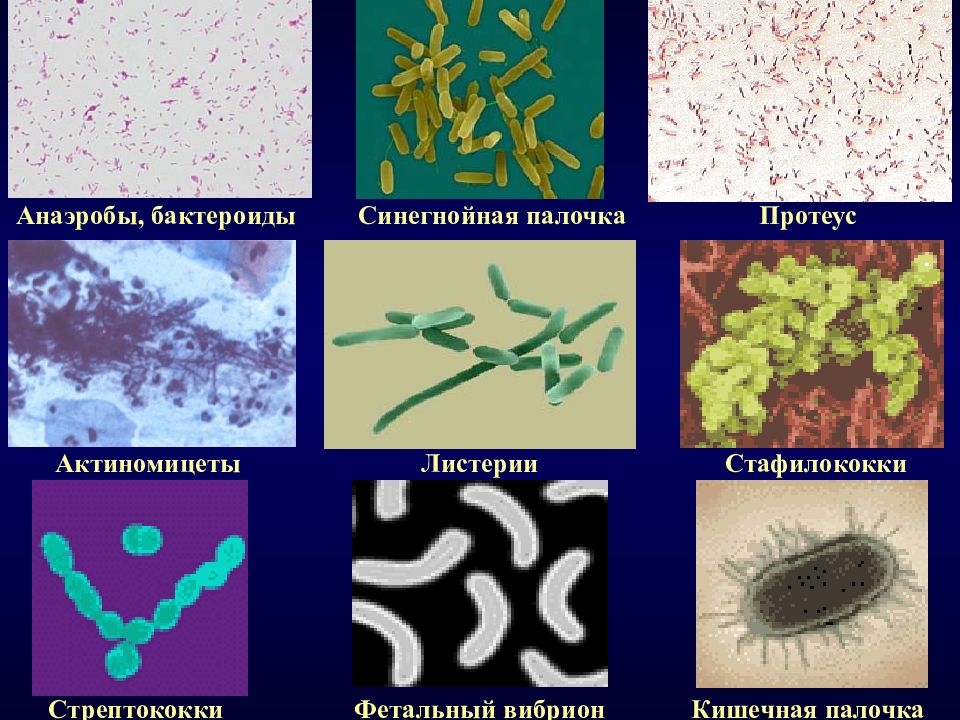 Бифидобактерии участвуют в производстве таких витаминов как В1, В2, В3
Бифидобактерии участвуют в производстве таких витаминов как В1, В2, В3
Причины снижения количества бифидобактерий
- Медикаментозное лечение (антибиотики, нестероидные противовоспалительные средства (НПВС) такие как анальгин, аспирин, слабительные средства)
- Неправильное питание (избыток жиров или белков или углеводов, голодание, неправильный режим питания, искусственное вскармливание)
- Кишечные инфекции (дизентерия, сальмонеллез, вирусные инфекции)
- Ферментопатии (целиакия, лактазная недостаточность)
- Хронические заболевания ЖКТ (хронический гастрит, панкреатит, холецистит, язвенная болезнь желудка или двенадцатиперстной кишки)
- Иммунные заболевания (иммунные дефициты, аллергии)
- Смена климатических зон
- Стресс
Лактобактерии
Норма лактобактерий
Дети до 1 года | 106 – 107 |
Дети старшего возраста | 107 – 108 |
Взрослые | 106 – 108 |
Лактобактерии занимают около 4-6% от общей массы бактерий кишечника. Лактобактерии являются не менее полезными, чем бифидобактерии. Их роль в организме следующая: поддержка уровня pH в кишечнике, производство большого количества веществ (молочная кислота, уксусная кислота, перекись водорода, лактоцидин, ацидофилин), которые активно используются для уничтожения патогенных микроорганизмов, а также вырабатывают лактазу.
Лактобактерии являются не менее полезными, чем бифидобактерии. Их роль в организме следующая: поддержка уровня pH в кишечнике, производство большого количества веществ (молочная кислота, уксусная кислота, перекись водорода, лактоцидин, ацидофилин), которые активно используются для уничтожения патогенных микроорганизмов, а также вырабатывают лактазу.
Причины снижения количества лактобактерий
- Медикаментозное лечение (антибиотики, нестероидные противовоспалительные средства (НПВС) такие как анальгин, аспирин, слабительные средства)
- Неправильное питание (избыток жиров или белков или углеводов, голодание, неправильный режим питания, искусственное вскармливание)
- Кишечные инфекции (дизентерия, сальмонеллез, вирусные инфекции)
- Хронические заболевания ЖКТ (хронический гастрит, панкреатит, холецистит, язвенная болезнь желудка или двенадцатиперстной кишки)
- Стресс
Эшерихии (E.coli типичные)
Норма эшерихий
Дети до 1 года | 107 – 108 |
Дети старшего возраста | 107 – 108 |
Взрослые | 107 – 108 |
Эшерихии появляются в организме человека с рождения и присутствуют в нем на протяжении всей жизни. Выполняют следующую роль в организме: участвуют в образовании витаминов группы В и витамина К, участвуют в переработке сахаров, вырабатывают антибиотикоподобные вещества (колицины) которые борются с патогенными организмами, усиливают иммунитет.
Выполняют следующую роль в организме: участвуют в образовании витаминов группы В и витамина К, участвуют в переработке сахаров, вырабатывают антибиотикоподобные вещества (колицины) которые борются с патогенными организмами, усиливают иммунитет.
Причины снижения количества эшерихий
- Гельминтозы
- Лечение антибиотиками
- Кишечные инфекции (дизентерия, сальмонеллез, вирусные инфекции)
Бактероиды
Норма бактероидов в кале
Дети до 1 года | 107 – 108 |
Дети старшего возраста | 107 – 108 |
Взрослые | 107 – 108 |
Бактероиды участвуют в пищеварении, а именно в переработке жиров в организме.
Причины увеличения содержания бактероидов
- Жировая диета (употребление большого количества жиров в пищу)
- Лечение антибиотиками
- Кишечные инфекции (дизентерия, сальмонеллез, вирусные инфекции)
Причины снижения содержания бактероидов
- Лечение антибиотиками
- Кишечные инфекции (дизентерия, сальмонеллез, вирусные инфекции)
Пептострептококки
Нормальное количество в кале
Дети до 1 года | 103 – 105 |
Дети старшего возраста | 105 – 106 |
Взрослые | 105 – 106 |
В норме пептострептококки живут в толстом кишечнике, при увеличении их количества и попадании в любую другую область нашего организма, они вызывают воспалительные заболевания. Участвуют в переработке углеводов и молочных белков. Вырабатывают водород, который в кишечнике превращается в перекись водорода и помогает контролировать рН в кишечнике.
Участвуют в переработке углеводов и молочных белков. Вырабатывают водород, который в кишечнике превращается в перекись водорода и помогает контролировать рН в кишечнике.
Причины увеличения содержания пептострептококков
- Употребление большого количества углеводов
- Кишечные инфекции
- Хронические заболевания ЖКТ
Энтерококки
Норма энтерококков
Дети до 1 года | 105 – 107 |
Дети старшего возраста | 105 – 108 |
Взрослые | 105 – 108 |
Энтерококки участвуют в переработке углеводов, в производстве витаминов, а также играют роль в создании местного иммунитета (в кишечнике). Количество энтерококков не должно превышать количество кишечных палочек, если их количество увеличивается, они могут вызвать ряд заболеваний.
Причины увеличения содержания энтерококков
- Снижение иммунитета, иммунные заболевания
- Пищевые аллергии
- Гельминтозы
- Лечение антибиотиками (в случае резистентности энтерококков к применяемому антибиотику)
- Неправильное питание
- Снижение количества кишечной палочки (эшерихии)
Стафилококки (сапрофитные стафилококки и патогенные стафилококки)
Норма сапрофитных стафилококков
Дети до 1 года | ≤104 |
Дети старшего возраста | ≤104 |
Взрослые | ≤104 |
Норма патогенных стафилококков
Дети до 1 года | — |
Дети старшего возраста | — |
Взрослые | — |
Стафилококки делятся на патогенные и непатогенные. К патогенным относятся: золотистый, гемолитический и плазмокоагулирующий, наиболее опасен золотистый. К непатогенным стафилококкам относятся негемолитический и эпидермальный.
К патогенным относятся: золотистый, гемолитический и плазмокоагулирующий, наиболее опасен золотистый. К непатогенным стафилококкам относятся негемолитический и эпидермальный.
Стафилококк не относится к нормальной микрофлоре кишечника, он попадает в организм из внешней среды вместе с пищей. Золотистый стафилококк, попадая в ЖКТ, обычно, вызывает токсические инфекции.
Причины появления стафилококка
Стафилококк может попасть в организм человека разными путями, начиная грязными руками, вместе с продуктами питания и заканчивая внутрибольничными инфекциями.
Клостридии
Норма клостридий
Дети до 1 года | ≤103 |
Дети старшего возраста | ≤105 |
Взрослые | ≤105 |
Клостридии участвуют в переработке белков, продуктом их переработки являются такие вещества как индол и скатол, которые по сути являются ядовитыми веществами, однако в небольших количествах эти вещества стимулируют перистальтику кишечника тем самым улучшая функцию эвакуации каловых масс. Однако при увеличении количества клостридий в кишечнике вырабатывается большее количество индола и скатола, что может привести к развитию такого заболевания как гнилостная диспепсия.
Однако при увеличении количества клостридий в кишечнике вырабатывается большее количество индола и скатола, что может привести к развитию такого заболевания как гнилостная диспепсия.
Причины увеличения количества клостридий
- Большое количество белка употребляемого в пищу
Кандида
Норма кандид
Дети до 1 года | ≤103 |
Дети старшего возраста | ≤104 |
Взрослые | ≤104 |
При увеличении количества кандид в кишечнике может развиться бродильная диспепсия, а также заметное увеличение количества кандид может спровоцировать развитие различных видов кандидоза.
Причины увеличения количества кандид
- Употребление большого количества углеводов в пищу
- Лечение антибиотиками (без применения в комплексе противогрибковых препаратов)
- Использование гормональных противозачаточных средств
- Беременность
- Сахарный диабет
- Стресс
Анализ кала на патогенную флору
Анализ кала на патогенную флору является тем же самым анализом кала на дисбактериоз. В бланке с результатами анализов он занимает место – Патогенные энтеробактерии. К группе патогенных энтеробактерий относят Сальмонеллу и Шигеллу как основных возбудителей инфекционных заболеваний кишечника.
В бланке с результатами анализов он занимает место – Патогенные энтеробактерии. К группе патогенных энтеробактерий относят Сальмонеллу и Шигеллу как основных возбудителей инфекционных заболеваний кишечника.
Патогенные (болезнетворные) бактерии (pathogenic bacteria, греч. pathos — страдание и genes — порождающий, рождающийся; греч. bacterion — палочка) — бактерии, паразитирующие на других организмах и способные вызывать инфекционные заболевания человек, среди которых особое место занимают кишечные инфекции. Кишечные инфекции — целая группа заразных заболеваний, которые в первую очередь повреждают пищеварительный тракт. Заражение происходит при попадании возбудителя инфекции через рот, как правило, при употреблении зараженных пищевых продуктов и воды. Всего таких заболеваний более 30, в частности, к ним относятся: холера, брюшной тиф, ботулизм, сальмонеллез, дизентерия и т.п. Среди кишечных заболеваний самое безобидное — это пищевое отравление. Возбудителями кишечных инфекций могут быть как сами бактерии (сальмонеллез, брюшной тиф, холера), так и их токсины (ботулизм). Некоторые бактерии могут вызывать язвенную болезнь и рак желудка, а также хронический гастрит.
Возбудителями кишечных инфекций могут быть как сами бактерии (сальмонеллез, брюшной тиф, холера), так и их токсины (ботулизм). Некоторые бактерии могут вызывать язвенную болезнь и рак желудка, а также хронический гастрит.
Сальмонелла
В норме сальмонеллы в результате анализа быть не должно!
Вызывает такое заболевание как сальмонеллез, которое проявляется сильным токсическим поражением кишечника. Основными переносчиками являются водоплавающие птицы.
Причины появления сальмонеллы
- Употребление плохо обработанного или сырого мяса
- Употребление плохо обработанных или сырых яиц
- Контакт с переносчиками
- Контакт с водой зараженной сальмонеллой
- Грязные руки
Шигелла
В норме шигеллы в результате анализа быть не должно!
Вызывает такое заболевание как дизентерия, которое также поражает кишечник и проявляется сильным токсическим поражением кишечника. Основными путями заражения являются молочные продукты, сырые овощи, зараженная вода, люди больные дизентерией.
Основными путями заражения являются молочные продукты, сырые овощи, зараженная вода, люди больные дизентерией.
Причины появления шигеллы
- Употребление или контакт с зараженной водой
- Употребление зараженных продуктов питания
- Контакт с людьми болеющими дизентерией
- Грязные руки и контакт с зараженными поверхностями (посуда, игрушки)
Часто задаваемые вопросы
Как правильно подготовиться к анализу?
Применение некоторых медикаментов может повлиять на результат анализа кала. Поэтому их применение необходимо приостановить или прекратить в период подготовки к сдаче анализа кала после консультации с вашим врачом.
- Отменить прием препаратов (после личной консультации Вашего лечащего врача). К препаратам, которые могут повлиять на результаты анализов кала, относятся следующие:
- Противодиарейные препараты (Смекта, Неосмектин, Полифан, Имодиум, Энтерол)
- Противогельминтные препараты (Немозол, Декарис, Вермокс, Гельминтокс)
- Антибиотики — любые виды
- Лечебные и очищающие клизмы
- Слабительные препараты (Бисакодил, Экстракт сенны, Форлакс, Порталак)
- НПВП – нестероидные противовоспалительные препараты (Аспирин, Парацетамол, Ибупрофен)
- Также нельзя использовать кал который был в контакте с чистящими или дезинфицирующими средствами, которые используются для чистки унитаза, водой или мочой.


Как правильно собрать кал для анализа?
- Помочитесь перед сбором материала, для того чтобы моча не попала в кал.
- Необходимо взять чистую, сухую ёмкость, куда будет проводиться дефекация.
- Из полученного материала необходимо взять 8-10 см3 (~2 чайные ложки). Кал забирается при помощи специальной «ложки» которая встроена в крышку специального контейнера, который вам должны выдать для сбора кала.
- Кал для анализа собирается из разных участков каловых масс (сверху, с боков, изнутри)
- Материал (кал) кладется в выданный вам контейнер, и плотно закрывается.
- Необходимо подписать контейнер (Ваше имя и фамилию, дату сбора анализа)
Как хранить кал перед отправкой в лабораторию?
Материал для анализа кала на дисбактериоз и на кишечную инфекцию необходимо доставить в лабораторию в максимально короткие сроки 30-40 минут (максимум 1,5 -2 часа). Чем больше времени прошло со времени сбора материала и момента доставки материала в лабораторию, тем менее достоверны будут анализы. Проблема в том, что большая часть бактерий кишечника являются анаэробными, то есть они живут в среде без кислорода, при контакте с ним умирают. Это может повлиять на достоверность результата. Поэтому хранить хоть какое либо время свыше рекомендуемых максимальных 2 часа, категорически не рекомендуется.
Проблема в том, что большая часть бактерий кишечника являются анаэробными, то есть они живут в среде без кислорода, при контакте с ним умирают. Это может повлиять на достоверность результата. Поэтому хранить хоть какое либо время свыше рекомендуемых максимальных 2 часа, категорически не рекомендуется.
Примечание
Полный (максимально подробный и точный) микробиологический анализ кала (анализ микробиоты толстой кишки) невозможно провести культурально зависимым способом, т.к. не все бактерии можно выделить и культивировать в искусственных условиях. В общем, проблема с микробиомом состоит в том, что мы можем выращивать в лаборатории менее 5% видов микробов, остальные просто слишком привередливы. Сегодня используются более технологичные культурально-независимые методы анализа:
Обзор популярных методов анализа микробиоты
Современные анализы кишечной микробиоты проводятся культурально-независимыми способами, среди которых можно выделить два наиболее известных на сегодняшний день в России метода — метод газовой хроматографии масс-спектрометрии микробных маркеров и метагеномный анализ микробиоты с помощью секвенирования нуклеиновых кислот. Обе методики имеют свои преимущества и недостатки (стоимость, время), включая погрешности в самих результатах и ограничения в использовании, а поэтому они совершенствуются до сих пор. Но несмотря на определенные недочеты, указанные технологии при грамотном использовании все же являются мощным инструментарием на пути к достижению целей персонализированнной медицины.
Обе методики имеют свои преимущества и недостатки (стоимость, время), включая погрешности в самих результатах и ограничения в использовании, а поэтому они совершенствуются до сих пор. Но несмотря на определенные недочеты, указанные технологии при грамотном использовании все же являются мощным инструментарием на пути к достижению целей персонализированнной медицины.
Информация к разделу: Дисбактериоз
См. отдельно: Типичный анализ микробиома кишечника по образцам стула (метод метагеномики)
Будьте здоровы!
ССЫЛКИ К РАЗДЕЛУ О ПРЕПАРАТАХ ПРОБИОТИКАХ
- ПРОБИОТИКИ
- ДОМАШНИЕ ЗАКВАСКИ
- БИФИКАРДИО
- КОНЦЕНТРАТ БИФИДОБАКТЕРИЙ ЖИДКИЙ
- ПРОПИОНИКС
- ЙОДПРОПИОНИКС
- СЕЛЕНПРОПИОНИКС
- МИКРОЭЛЕМЕНТНЫЙ СОСТАВ
- ПРОБИОТИКИ С ПНЖК
- БИФИДОБАКТЕРИИ
- ПРОПИОНОВОКИСЛЫЕ БАКТЕРИИ
- ПРОБИОТИКИ И ПРЕБИОТИКИ
- СИНБИОТИКИ
- РОЛЬ МИКРОБИОМА В ТЕРАПИИ РАКА
- АНТИОКСИДАНТНЫЕ СВОЙСТВА
- АНТИОКСИДАНТНЫЕ ФЕРМЕНТЫ
- АНТИМУТАГЕННАЯ АКТИВНОСТЬ
- МИКРОФЛОРА КИШЕЧНОГО ТРАКТА
- МИКРОБИОМ ЧЕЛОВЕКА
- МИКРОФЛОРА И ФУНКЦИИ МОЗГА
- ПРОБИОТИКИ И ХОЛЕСТЕРИН
- ПРОБИОТИКИ ПРОТИВ ОЖИРЕНИЯ
- МИКРОФЛОРА И САХАРНЫЙ ДИАБЕТ
- ПРОБИОТИКИ и ИММУНИТЕТ
- МИКРОБИОМ И АУТОИММУННЫЕ БОЛЕЗНИ
- ПРОБИОТИКИ и ГРУДНЫЕ ДЕТИ
- ПРОБИОТИКИ, БЕРЕМЕННОСТЬ, РОДЫ
- ДИСБАКТЕРИОЗ
- ВИТАМИННЫЙ СИНТЕЗ
- АМИНОКИСЛОТНЫЙ СИНТЕЗ
- АНТИМИКРОБНЫЕ СВОЙСТВА
- СИНТЕЗ ЛЕТУЧИХ ЖИРНЫХ КИСЛОТ
- СИНТЕЗ БАКТЕРИОЦИНОВ
- ФУНКЦИОНАЛЬНОЕ ПИТАНИЕ
- АЛИМЕНТАРНЫЕ ЗАБОЛЕВАНИЯ
- ПРОБИОТИКИ ДЛЯ СПОРТСМЕНОВ
- ПРОИЗВОДСТВО ПРОБИОТИКОВ
- ЗАКВАСКИ ДЛЯ ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ
- НОВОСТИ
бактероиды снижены лечение — 16 рекомендаций на Babyblog.
 ru
ruСреди ВЗОМТ важная роль отводится хроническому эндометриту (ХЭ). По данным разных авторов, распространенность ХЭ варьирует от 10% до 85%], что обусловлено определенными трудностями диагностики, клинической и морфологической верификации этого заболевания. Частота встречаемости ХЭ имеет тенденцию к неуклонному росту, что связано с широким использованием внутриматочных средств контрацепции, а также с ростом числа абортов и различных внутриматочных вмешательств. В 80-90% случаев ХЭ выявляется у женщин детородного возраста (средний возраст заболевших — 36 лет) и обуславливает у них нарушения менструальной и репродуктивной функций, приводя, в конечном итоге, к развитию бесплодия, неудачам в программах экстракорпорального оплодотворения (ЭКО) и переноса эмбриона (ПЭ), невынашиванию беременности и осложненному течению гестационного процесса и родов.
Большую роль в развитии ХЭ играют инфекционные агенты. На сегодняшний день ХЭ рассматривается как клинико-морфологический синдром, при котором вследствие персистирующего повреждения эндометрия инфекционным агентом возникают множественные вторичные морфологические и функциональные изменения, нарушающие циклическую биотрансформацию и рецептивность слизистой оболочки тела матки.
В 95% случаев ХЭ является первичным, развиваясь непосредственно в эндометрии за счет внедрения экзогенных штаммов микроорганизмов, передающихся половым путем, или размножения условно-патогенной микрофлоры в эндометрии после внутриматочных лечебных и диагностических манипуляций. Лишь в 5% случаев эндометрит носит вторичный характер, развиваясь при попадании инфекции в эндометрий из экстрагенитальных очагов гематогенным, лимфогенным или нисходящим путями.
Согласно современным представлениям, факторами риска развития ХЭ являются инвазивные манипуляции в полости матки (аборты, гистероскопия, диагностическое выскабливание, биопсия эндометрия, гистеросальпингография, введение внутриматочных контрацептивов, внутриматочная инсеминация, ЭКО и др.), послеродовые инфекционно-воспалительные осложнения, заболевания мочевыводящей системы, хронический сальпингоофорит в анамнезе, наличие какой-либо иной гинекологической патологии (миома матки, гипер- и гипопластические процессы эндометрия, полипы эндометрия и цервикального канала, чаще всего, железисто-фиброзные), а также перенесенные операции на органах малого таза.
В основе современной классификации ХЭ лежат различные этиологические факторы этого заболевания, с учетом которых принято выделять неспецифический и специфический ХЭ. В первом случае специфическая микрофлора в клетках слизистой оболочки тела матки не выявляется, а к развитию неспецифического воспаления в эндометрии предрасполагают длительная внутриматочная контрацепция, длительно нелеченый вялотекущий бактериальный вагиноз (восходящий путь инфицирования эндометрия), лучевая терапия органов малого таза, а также ВИЧ-инфекция. Специфический ХЭ может быть хламидийной, вирусной (вирус простого герпеса — ВПГ, цитомегаловирус — ЦМВ, энтеровирусы), бактериальной (возбудители туберкулеза, гонореи, менингита, сифилиса), микоплазменной, грибковой, протозойной (токсоплазма, шистосома) и паразитарной этиологии, а также может развиваться на фоне саркоидоза. По данным В. И. Краснопольского и соавт., спектр генитальной инфекции, выявленной при ПЦР-диагностике отделяемого из цервикального канала и полости матки у пациенток с ХЭ, включает в себя: хламидиоз — в 14,9% случаев, генитальный герпес — в 33,6%, уреаплазмоз — в 37,8%, микоплазмоз — в 11,6%, ЦМВ — в 18,9% случаев.
Важная роль в развитии ХЭ на сегодняшний день принадлежит облигатно-анаэробным микроорганизмам — бактероидам и пептострептококкам — в сочетании с микроаэрофилами — микоплазмами и гарднереллами, а также факультативно-анаэробным микроорганизмам, таким как эшерихии, энтерококки, стрептококки группы В. Так, при невынашивании беременности, ассоциированной с ХЭ, вирусно-бактериальная контаминация эндометрия выявляется у 55,8% женщин, тогда как чистая бактериальная обсемененность эндометрия верифицируется только у 12,3% пациенток, а моновирусная контаминация эндометрия, преимущественно ВПГ или ЦМВ, — у 31,9% женщин.
Среди персистирующих вирусов в генезе ХЭ наибольшее значение имеют герпес-вирусные (ВПГ, Herpes zoster и ЦМВ), энтеровирусные (вирусы Коксаки А и В) и аденовирусные инфекции. По данным Л. С. Лазоревской и соавт., вирусы Коксаки А выявляются у 98%, Коксаки В — у 74,5%, энтеровирусы — у 47,1%, ЦМВ — у 60,8% и ВПГ — у 56,9% женщин с привычным невынашиванием беременности (ПНБ).
К основным клиническим симптомам, выявляемым у пациенток с ХЭ, относятся маточные кровотечения циклического характера и/или перименструальные кровянистые выделения из половых путей, болевой синдром, диспареуния, серозные и гноевидные выделения из половых путей, значительно снижающие качество жизни женщин с этим заболеванием. Кроме того, длительное мало- или асимптомное течение ХЭ может привести к нарушению репродуктивной функции в виде ПНБ и бесплодия. Важно отметить, что при изолированном ХЭ без сочетанной гинекологической патологии в клинической картине заболевания преобладают нарушения менструального цикла и периодические тянущие боли в низу живота. С другой стороны, при сочетании ХЭ с простой типичной гиперплазией эндометрия ведущими в клинической картине симптомами являются нарушения менструальной функции и невынашивание беременности, а у пациенток с ХЭ и железисто-фиброзным полипом эндометрия наиболее часто встречаются кровотечения по типу метроррагии и болевой синдром.
Болевой синдром при ХЭ выявляется у 33-89% женщин в виде периодических, реже постоянных, тянущих болей внизу живота и диспареунии. Учитывая современные особенности течения ХЭ с преобладанием стертых форм заболевания, в последнее время все чаще основным клиническим признаком ХЭ является нарушение репродуктивной функции в виде бесплодия, включая неэффективность ЭКО, и ПНБ. Л. Н. Кузьмичев указывает на корреляцию ХЭ с высокой частотой неудач программ ЭКО и репродуктивных потерь в циклах вспомогательных репродуктивных технологий (ВРТ). По данным разных авторов, частота ХЭ у женщин с бесплодием колеблется от 12,3% до 60,4%, являясь единственной причиной бесплодия в 18,8% случаев. Соотношение первичного бесплодия к вторичному у женщин с гистологически подтвержденным ХЭ составляет от 1:1,5 до 1:2,4 случая. У пациенток с несколькими неудачными попытками ЭКО в анамнезе частота выявления ХЭ достигает 80%, причем среднее количество неудач в программах ВРТ равно 3,1 ± 0,4 на одну женщину. По данным Г. Т. Сухих и А. В. Шуршалиной , ХЭ является единственной верифицированной причиной невынашивания беременности в 47,4-52,1% случаев , тогда как при ПНБ, по данным разных авторов, частота выявления ХЭ колеблется в диапазоне от 33% до 86,7% случаев вне зависимости от клинической картины прерывания беременности.
На начальных этапах течения ХЭ за счет наличия определенного компенсаторного механизма параметры рецептивности эндометрия сохраняются на достаточном для реализации репродуктивных задач уровне. Эффективность функционирования этого механизма определяется активностью иммунной системы, адекватностью стероидной регуляции, наличием сочетанной гинекологической патологии, а также характеристиками самого повреждающего агента. Длительная персистенция микробных агентов в эндометрии вызывает изменение антигенной структуры инфицированных клеток как за счет собственно инфекционных антигенов, включаемых в структуру поверхностных мембран, так и за счет образования новых клеточных антигенов, детерминированных клеточным геномом. В результате развивается иммунный ответ на гетерогенизированные аутоантигены по типу реакции гиперчувствительности замедленного типа, приводя к появлению аутоантител и развитию хронической формы ДВС-синдрома. Последнее вызывает возникновение локальных микротромбозов и инфарктов в области плацентации с последующей отслойкой плодного яйца/плаценты, что в совокупности с повреждающим действием самого инфекционного агента, гормональным дисбалансом и воздействием аутоантител приводит к срыву механизмов локальной резистентности, что клинически проявляется в виде патологии репродукции.
Нарушение механизмов локальной резистентности способствует персистенции микробных агентов в эндометрии, инициируя хроническое течение воспалительного процесса.
Таким образом, причинами хронического течения (хронизации) воспалительного процесса в эндометрии являются:
отсутствие своевременной диагностики и лечения;
длительная персистенция инфекционного агента;
длительная стимуляция иммунных клеток;
незавершенность заключительной фазы воспаления;
эволюция микробных факторов, с преобладанием доли вирусной и условно-патогенной микрофлоры;
хронический эндоцервицит;
нарушения тканевого гомеостаза;
повышенная регенерация ткани.
Важно подчеркнуть, что длительное течение ХЭ сопряжено с развитием вторичных морфофункциональных изменений эндометрия, заключающихся в активации склеротических процессов с повреждением экстрацеллюлярного матрикса, нарушением медиаторных межклеточных взаимодействий, изменением ангиоархитектоники ткани и развитием ее ишемии. При этом в большинстве случаев при гистологическом исследовании эндометрия выявляется «неполноценная морфологическая картина ХЭ», а вследствие нарушенного иммунного барьера со временем часто происходит смена ведущего микробного агента в сторону преобладания условно-патогенной микрофлоры. Подобные особенности создают определенные трудности в решении вопроса о необходимости назначения антибактериальной терапии, особенно в тех случаях, когда выявление патогена в полости матки затруднительно.
Остается актуальной и требует дальнейшего совершенствования с учетом современных знаний и возможностей проблема научно обоснованной терапии заболевания. Лечение ХЭ должно быть комплексным, этиологически и патогенетически обоснованным, поэтапным и базироваться на результатах максимально тщательного и точного обследования состояния эндометрия. Большинство ученых сходятся во мнении о целесообразности проведения комплексной терапии ХЭ в два этапа.
Сущность первого этапа заключается в элиминации повреждающего эндометрий микробного фактора и/или снижении активности вирусной инвазии посредством проведения этиотропной терапии антибиотиками широкого спектра действия, анаэробными средствами и/или противовирусными препаратами. При стерильных посевах эндометрия или невозможности проведения микробиологического исследования у пациенток с признаками ХЭ допустимо проведение эмпирической антибактериальной и противовирусной терапии препаратами широкого спектра действия со сменой групп препаратов в течение нескольких курсов в случае необходимости.
Цель второго этапа лечения ХЭ заключается в восстановлении морфофункционального потенциала эндометрия путем устранения результатов вторичных повреждений ткани — коррекции фиброзирующих и склеротических процессов, последствий ишемии, восстановлении гемодинамики и активности рецепторного аппарата эндометрия. Характер терапии ХЭ на данном этапе зависит от морфологического состояния эндометрия и наличия сочетанной гинекологической патологии. Первый аспект сводится к выраженности воспалительной реакции и наличию обширных участков склероза и/или атрофии в эндометрии, что определяется длительностью течения ХЭ и приводит к нарушению рецептивности эндометрия при продолжительности заболевания более 2 лет.Несмотря на разнообразие предлагаемых вариантов фармакологической коррекции, лечение ХЭ сопряжено со значительными методическими и практическими трудностями. ХЭ характеризуется нарушением ангиоархитектоники эндометрия, в частности, имеет место склерозирование стенок сосудов и образование периваскулярного склероза вокруг спиральных артерий и в базальных отделах эндометрия, что приводит к развитию ишемии эндометрия. По данным ряда авторов, в патогенезе нарушения ангиоархитектоники при ХЭ имеют значение хроническое воспаление с повреждающим действием воспалительного инфильтрата, патологическая регенерация и склероз, связанный с действием провоспалительных цитокинов.
Одним из грозных осложнений ХЭ является синдром Ашермана. Синдром Ашермана — комплекс симптомов нарушений менструальной и детородной функций, обусловленных внутриматочными синехиями. Внутриматочные синехии образуются вследствие травматизации слизистой оболочки матки, чаще при лечебно-диагностических выскабливаний стенок полости матки, аборте, особенно на фоне ХЭ. Выраженный спаечный процесс приводит не только к деформации полости матки, а иногда облитерации всей полости, при этом эндометрий подвергается атрофическим изменениям и, как следствие, развиваются вторичные аменорея и бесплодие. Появление вышеуказанных симптомов у женщин, перенесших внутриматочное вмешательство, позволяет предположить синдром Ашермана, для которого характерна отрицательная проба с эстрогенами и прогестероном — отсутствие менструальноподобного кровотечения. Эффективным диагностическим методом исследования является гистероскопия, позволяющая четко визуализировать внутриматочные синехии, деформацию или облитерацию полости матки.
Лечение больных с синдромом Ашермана оперативное, проводят рассечение синехий под контролем гистероскопа с последующим введением в полость матки внутриматочного контрацептива, далее назначается антифиброзирующая и циклическая гормональная терапия с тем, чтобы подавить патологический рост соединительной ткани и провести стимуляцию функциональной активности эндометрия. После 3-6 месяцев консервативной терапии внутриматочный контрацептив удаляется.
С целью повышения клинической эффективности комплексной терапии ХЭ необходимо четкое понимание механизмов нарушения иммунного гомеостаза, патологического роста соединительной ткани, фиброзирования и склерозирования при хроническом воспалительном процессе эндометрия с целью осуществления терапии, направленной на коррекцию указанных процессов, являющихся одним из ключевых звеньев патогенетической цепи, приводящих к нарушению репродуктивной функции при ХЭ.
Главные компоненты внеклеточного матрикса рыхлой соединительной ткани — протеогликаны, гликопротеиды, волокна соединительной ткани и другие гликоконъюгаты. Наиболее широко представленным компонентом в составе внеклеточного матрикса соединительной ткани из гликоконъюгатов является гиалуроновая кислота, которая синтезируется в основном фибробластами.
Гиалуроновая кислота — несульфированный гликозаминогликан, входящий в состав соединительной ткани и являщийся одним из основных компонентов внеклеточного матрикса. Гиалуроновая кислота деградируется семейством ферментов, называемых гиалуронидазами — термин был впервые предложен еще в 1940 г. Karl Meyer. В организме человека существуют, по меньшей мере, семь типов гиалуронидазоподобных ферментов, некоторые из которых являются супрессорами опухолеобразования.
Гиалуронидазы обладают способностью увеличивать проницаемость тканей за счет снижения вязкости мукополисахаридов, входящих в их состав, так, например, тестикулярная гиалуронидаза, содержащаяся в сперматозоидах, способствует процессу оплодотворения яйцеклетки.
Следует отметить, что компенсаторное повышение активности гиалуронидазы происходит при воспалении, отеке, а потеря активности гиалуронидазы приводит к накоплению гиалуроновой кислоты, что в свою очередь приводит к фиброзу и склерозу. Фиброз представляет собой универсальный процесс, основу которого составляет накопление протеинов внеклеточного матрикса и уплотнение соединительной ткани с появлением рубцовых изменений, возникающее, как правило, в результате хронического воспаления, а склероз — замена паренхимы органов плотной соединительной тканью, то есть уплотнение органов, вызванное гибелью функциональных элементов и заменой их фиброзной тканью. Среди наиболее распространенных причин патологического фиброза и склероза главное место занимают воспалительные процессы, особенно те, которые связаны с хроническим воспалительным процессом, обусловленным как инфекционными, так и неинфекционными факторами.
Ключевым механизмом формирования продуктивной фазы воспаления является стимуляция фибробластов, как продуктами деструкции соединительной ткани, так и цитокинами, синтезируемыми макрофагами и другими продуктами деструкции соединительной ткани.
Хронический воспалительный процесс приводит к нарушению функции клеток и межклеточного вещества соединительной ткани и в итоге гиперплазии (патологический рост) соединительной ткани.
Следует отметить, что воспаление, завершающееся гиперплазией соединительной ткани, всегда протекает на фоне нарушения функций иммунной системы — ослабления или гиперактивности иммунного ответа. При гиперактивности иммунной системы активированные фагоциты усиленно продуцируют провоспалительные цитокины — мощные стимуляторы функции фибробластов. Фагоцитирующие клетки «выбрасывают» в окружающую среду и свободные радикалы, которые, уничтожая инфекцию, повреждают также и нормальные ткани. В случае ослабленной иммунной системы развивающееся хроническое воспаление также сопровождается повреждением тканей, что активирует продуктивную фазу воспаления и ведет к развитию патологического роста соединительной ткани.
Вызванный иммунными нарушениями системный воспалительный процесс составляет общий и главный стержень диффузных болезней соединительной ткани.
Большую роль в воспалении играет процесс оксиадтивного стресса и перекисного окисления липидов, возникающих с самого начала и сопровождающих постоянно воспалительный процесс. Мембраны иммунокомпетентных клеток повреждаются при активации перекисных процессов в первую очередь. В частности, лимфоциты, включенные в воспалительный процесс, меняют свои функции и начинают активно продуцировать фактор роста фибробластов, благодаря которому происходит усиленная пролиферация фибробластов, активируется продукция коллагена (Серов В. В. и соавт.).
Таким образом, становится очевидным, что подавление фиброзообразования — одна из первоочередных задач терапии фиброзирующих процессов при хроническом воспалительном процессе эндометрия, помимо коррекции иммунитета и микроциркуляции. Коррекция иммунитета и микроциркуляции, усиление антиоксидантной защиты, в свою очередь, также оказывают непрямой антифиброзирующий эффект. Антиоксиданты способны приводить к торможению активации и пролиферации фибробластов, снижая активность реакции пероксидации в клеточных мембранах, уменьшая накопление фибриллярных коллагенов 1-го и 3-го типа и протеинов внеклеточного матрикса.
Среди прямых антифибринозных средств основными ферментными препаратами являются коллагеназа и гиалуронидаза, активность которых может быть подавлена тканевыми ингибиторами металлопротеиназ. Гиалуронидаза является основным ферментом, гидролизующим крупномолекулярные компоненты внеклеточного матрикса, включая не только гиалуроновую кислоту, но и другие крупные молекулы внеклеточного матрикса соединительной ткани.
Анализ данных литературы подтверждает, что наиболее распространенным способом воздействия на соединительную ткань с целью предупреждения ее гиперплазии является деполимеризация внеклеточного матрикса ферментами типа гиалуронидазы.
Одной из главных причин низкой эффективности при назначении гиалуронидазы является наличие в организме большого количества ингибиторов фермента. Филогенетически в организме выработалось большое количество ингибиторов гиалуронидазы. Продукты гидролиза матрикса (протеогликаны и гликозаминогликаны) и структурных элементов соединительной ткани — коллагеновых волокон — являются стимуляторами синтеза этих же соединений.
В связи с тем, что гиалуронидаза (препараты) имеют ряд серьезных недостатков, а именно: реактогенность, термолабильность, быструю инактивацию в крови многочисленными ингибиторами, необходимость назначения длительными курсами, развитие аллергических реакций, назрела необходимость создания препарата, лишенного указанных недостатков.
Решение проблемы стабилизации фермента, блокирования ингибиторов гиалуронидазы и развивающейся воспалительной реакции было найдено А. В. Некрасовым путем создания полифункционального фармакологического средства Лонгидазы, представляющего собой конъюгат гиалуронидазы с высокополимерным носителем — азоксимера бромидом (сополимер N-оксида 1,4-этиленпиперазина и (N-карбоксиметил)-1,4-этиленпиперазиний бромида). Благодаря химической связи гиалуронидазы с азоксимера бромидом препарат становится защищенным от действия ингибиторов, образующихся в воспалительном очаге, в течение длительного времени сохраняется его ферментативная активность, уменьшается число побочных реакций при ведении Лонгидазы.
Лонгидаза по механизму действия существенно отличается от всех аналогичных препаратов, имеющих в своей основе гиалуронидазу. Эффективность Лонгидазы на всех стадиях, начиная от воздействия на организм повреждающего фактора и заканчивая стадией развития фиброза, доказана экспериментальным путем (Иванова А. С. и соавт.). Лонгидаза инактивирует цитотоксические свойства химических агентов, защищая тем самым клетки от повреждения. Положительное действие Лонгидазы на этом этапе заключается в следующем:
прямая инактивация активных форм кислорода и других свободных радикалов, повреждающих клетки и ткани;
хелатирование (связывание) и удаление из очага воспаления активных ионов железа, самых мощных стимуляторов радикальных реакций;
снижение избыточного синтеза провоспалительных цитокинов, прежде всего ФНО-альфа.
Описанные свойства Лонгидазы обуславливают ее высокий терапевтический эффект, способность препарата не только тормозить развитие продуктивной фазы воспаления, но и вызвать обратное развитие сформировавшейся патологической соединительной ткани (фиброза, спаек, склеротических образований).
Важно отметить, что Лонгидаза не повреждает нормальную соединительную ткань, а вызывает деструкцию измененной по составу и структуре патологической соединительной ткани. Лонгидаза не обладает антигенными свойствами, митогенной, поликлональной активностью, не оказывает аллергизирующего, мутагенного, эмбриотоксического, тератогенного и канцерогенного действия.
При клиническом применении Лонгидазы не отмечено местных и системных пробочных реакций.
Лонгидаза рекомендуется к применению в составе комплексной терапии для лечения заболеваний, сопровождающихся ростом соединительной ткани, для воздействия на гранулематозно-фиброзный процесс. Рекомендуемые дозировки 3000 МЕ суппозитории для вагинального и ректального применения курсом от 10 до 20 введений. Схема ведения корригируется в зависимости от тяжести, стадии и длительности заболевания: свечи вводятся через день или с перерывами в 2-3 дня.
Лонгидаза имеет выраженные преимущества по сравнению с ферментативным аналогом — препаратами на основе гиалуронидазы.
Полученные данные свидетельствуют о том, что Лонгидаза является принципиально новым полифункциональным фармакологическим средством, способным подавлять острую фазу воспаления, останавливать реактивный рост соединительной ткани и вызывать обратное развитие фиброза, то есть проявляет целый комплекс фармакологических свойств, направленных на коррекцию сложнейших ауторегуляторных реакций соединительной ткани.
Как известно, транспорт антибактериальных препаратов и других лекарственных средств в эндометрий при его хроническом воспалении значительно затруднен, прежде всего, вследствие гиперплазии соединительной ткани, а также гемодинамических и микроциркуляторных нарушений в эндометрии (венозный стаз). Увеличение биодоступности лекарственных средств, в частности при ХЭ, возможно при применении протеолитических ферментов, в частности гиалуронидаз.
Сочетание приема антибиотиков с препаратом Лонгидаза в комплексной лечении больных с ХЭ приводит к значительному повышению клинической эффективности проводимой терапии, вследствие как увеличения биодоступности антибактериальных средств, так и нивелирования фиброза и склероза эндометрия, которые являются основными причинами нарушения репродуктивной функции у данной категории пациенток.
Таким образом, проведение комплексной патогенетически обоснованной терапии ХЭ способствует повышению клинической эффективности лечения. К критериям эффективности лечебных мероприятий при ХЭ относятся: полное купирование клинических симптомов заболевания в сочетании с элиминацией патогенной микрофлоры из полости матки на фоне нормализации в эндометрии уровней иммунокомпетентных клеток, провоспалительных цитокинов; восстановление микроциркуляции эндометрия; улучшение реологических свойств крови; снижения интенсивности процессов фиброзирования, склерозирования и стабилизации компонентов экстрацеллюлярного матрикса. Заключительным успехом лечения ХЭ является восстановление репродуктивной функции с последующим наступлением беременности и нормальным течением гестационного процесса.
Хронический эндометрит — проблема и решения
По данным различных авторов, факторами риска развития ХЭ являются все инвазивные вмешательства в полости матки (гистероскопия, выскабливание полости матки, биопсия эндометрия, гистеросальпингография, манипуляции в программах вспомогательных репродуктивных технологий и др.), инфекционно-воспалительные осложнения после беременностей и родов, использование внутриматочных спиралей (ВМС), инфекции влагалища и шейки матки, бактериальный вагиноз, деформации полости матки с нарушением циклического отторжения эндометрия, лучевая терапия в области органов малого таза.
В современных условиях эндометрит характеризуется рядом особенностей: изменением этиологической структуры с увеличением значимости вирусной и условно-патогенной флоры, ростом резистентности флоры к фармакотерапии, трансформацией клинической симптоматики в сторону стертых форм и атипичного течения, длительными сроками терапии и высокой ее стоимостью. Современная общепризнанная классификация ХЭ, выстроенная по этиологическому фактору, претерпела в последнее десятилетие значительные изменения в сторону расширения перечня специфических форм заболевания. Существуют вирусные (вирус папилломы человека — ВПЧ, цитомегаловирус, вирус иммунодефицита человека), хламидийные, микоплазменные, грибковые, протозойные и паразитарные специфические эндометриты, отдельно выделен саркоидоз эндометрия . Для идентификации инфекционного агента обязательно проводить тщательное микробиологическое исследование соскоба эндометрия с определением чувствительности флоры к антибактериальным препаратам. В одной трети случаев при гистологически верифицированном ХЭ выявляются стерильные посевы эндометрия, что может свидетельствовать о важной роли условно-патогенной флоры в развитии воспалительного процесса или недостаточной детекции возбудителя, особенно в случае вирусной инвазии. Длительная и часто бессимптомная персистенция инфекционных агентов в эндометрии приводит к выраженным изменениям в структуре ткани, препятствуя нормальной имплантации и плацентации, формируя патологический ответ на беременность, а также вызывает нарушение пролиферации и нормальной циклической трансформации ткани. По различным данным, у больных с бесплодием частота ХЭ составляет в среднем 10% (от 7,8 до 15,4%). Среди женщин с верифицированным ХЭ в 60% случаев диагностируется бесплодие (в 22,1% — первичное бесплодие, в 36,5% — вторичное бесплодие), неудачные попытки ЭКО и переноса эмбрионов в анамнезе отмечены у 40% женщин. Клиническая картина ХЭ в значительной степени отражает глубину структурных и функциональных изменений в ткани эндометрия. Неполноценная циклическая трансформация эндометрия на фоне воспалительного процесса, нарушение процессов десквамации и регенерации функционального слоя эндометрия приводят к появлению основного симптома заболевания — маточных кровотечений (перименструальные, межменструальные). Достаточно постоянными являются жалобы на тянущие боли внизу живота, дисменорею и диспареунию, нередко отмечаются серозные и серозно-гноевидные бели. Среди клинических симптомов особое место занимает бесплодие (преимущественно вторичное), неудачные попытки ЭКО и НБ. Более половины пациенток в анамнезе имеют инвазивные вмешательства в полости матки и на органах малого таза. Часто хронический эндометрит сочетается с хроническим сальпингоофоритом и спаечным процессом в малом тазе. Важно отметить, что использование минимальных критериев для диагностики воспалительных заболеваний органов малого таза (ВЗОМТ), предложенных CDC (Centers for Disease Control and Prevention, США, 1993), информативно в случае с хроническим эндометритом только в 33% . В целом у 35-40% пациенток какие-либо клинические симптомы заболевания отсутствуют. Диагностика ХЭ основана на анализе клинических симптомов, данных анамнеза, эхографической картины. Обязательным этапом диагностики заболевания является морфологическое исследование эндометрия .
Эхографические критерии ХЭ впервые разработаны профессором В. Н. Демидовым с соавт. (1993 г.). Исследование проводят дважды на 5-7 и 17-21 день менструального цикла. Часто при ХЭ полость матки после менструации расширена до 0,4-0,7 см, в ряде случаев имеется асимметричность расширения полости матки и неровность внутренней поверхности эндометрия. Изменение структуры эндометрия также выражается в возникновении в зоне срединного М-эха участков повышенной эхогенности различной величины и формы. Внутри участков выявляются отдельные зоны неправильной формы и сниженной эхогенности. Другой важный признак — наличие пузырьков газа, иногда с характерным акустическим эффектом «хвоста кометы». В базальном слое эндометрия часто визуализируются четкие гиперэхогенные образования диаметром до 0,1-0,2 см, представляющие собой очаги фиброза, склероза или кальциноза. Довольно редким, но важным признаком эндометрита является асимметрия толщины передней и задней стенок эндометрия. О наличии патологии свидетельствует также истончение М-эха во второй трети менструального цикла менее 0,6 см. У каждой пятой больной отмечается наличие нескольких из перечисленных признаков .
В настоящее время для диагностики внутриматочной патологии широко используется гистероскопия. Наиболее частыми признаками воспалительного процесса в эндометрии являются: неравномерная толщина эндометрия — 30% случаев, полиповидные нарастания — 32,2%, неравномерная окраска и гиперемия слизистой оболочки — 23% и 11,8%, точечные кровоизлияния — 9%, очаговая гипертрофия слизистой оболочки — 7%. Трудности гистероскопической интерпретации данных связаны с отсутствием типичных макроскопических признаков хронического эндометрита, с очаговым характером воспалительного процесса и стертыми формами заболевания. По разным данным гистероскопия позволяет по макроскопическим признакам точно идентифицировать хронический эндометрит только в 16-35% случаев. При подозрении на ХЭ гистероскопия важна для исключения всего спектра внутриматочной патологии, но для верификации диагноза во всех случаях необходимо проведение морфологического исследования эндометрия.
«Золотым стандартом» диагностики ХЭ является морфологическое исследование эндометрия, которое должно быть обязательным звеном алгоритма обследования. Диагностическое выскабливание слизистой оболочки матки производят в среднюю и позднюю фазу пролиферации, на 7-10 день менструального цикла. В последние годы общепринятыми критериями морфологической диагностики ХЭ являются:
Воспалительные инфильтраты, состоящие преимущественно из лимфоидных элементов, расположенные чаще вокруг желез и кровеносных сосудов, реже диффузно. Очаговые инфильтраты имеют вид «лимфоидных фолликулов» и располагаются не только в базальном, но и во всех отделах функционального слоя, в состав их входят также лейкоциты и гистиоциты.
Наличие плазматических клеток.
Очаговый фиброз стромы, возникающий при длительном течении хронического воспаления, иногда захватывающий обширные участки.
Склеротические изменения стенок спиральных артерий эндометрия, появляющиеся при наиболее длительном и упорном течении заболевания и выраженной клинической симптоматике. Различия в трактовке гистологических особенностей ХЭ обусловлены наличием вариантов, которые определяются особенностями общей и тканевой реактивности, этиологическим фактором, продолжительностью заболевания, наличием обострений и степенью их выраженности.
Несмотря на значительное развитие фармакологической основы терапии, лечение ХЭ в настоящее время представляет значительные трудности для клиницистов.
Комплексная терапия ХЭ должна быть этиологически и патогенетически обоснованной, поэтапной и базироваться на результатах максимально тщательного обследования. Алгоритм лечения должен учитывать все звенья патогенеза заболевания.
Необходимость использования этиотропных препаратов для элиминирования повреждающего агента или снижение вирусной активности не вызывает сомнения. Принимая во внимание, что почти в 70% случаев ВЗОМТ применение рутинных методов диагностики не позволяет выявить этиологический фактор, эмпирическая антимикробная терапия при ХЭ всегда оправдана и приводит к достоверному уменьшению частоты клинических симптомов. При этом она должна обеспечивать элиминацию широкого спектра возможных патогенных микроорганизмов, включая хламидии, гонококки, трихомонады, грамотрицательные факультативные бактерии, анаэробы, стрептококки и вирусы. Оправдано назначение нескольких курсов этиотропной терапии со сменой групп препаратов.
Выбор группы лекарственных препаратов зависит от данных микробиологического исследования содержимого влагалища и полости матки, анамнестических указаний на использование антибактериальных препаратов в течение последнего года, от выраженности клинической симптоматики и обширности воспалительного процесса.
Основу базовой антибактериальной терапии должны составлять комбинации фторхинолонов и нитроимидазолов; цефалоспоринов III поколения, макролидов и нитроимидазолов, а также сочетание защищенных пенициллинов (амоксициллин/клавулановая кислота) с макролидами. При наличии вирусной инфекции (чаще вируса простого герпеса) в зависимости от клинической картины применяют современные аналоги нуклеозидов (Ацикловир или Валтрекс), как правило, в длительном режиме — в течение не менее 3-4 месяцев.
Одновременно на основании данных иммунного и интерферонового статуса проводится коррекция иммунных нарушений.
Второй этап лечения ХЭ направлен на восстановление морфофункционального потенциала ткани и устранение последствий вторичных повреждений: коррекция метаболических нарушений и последствий ишемии, восстановление гемодинамики и активности рецепторного аппарата эндометрия. Данный этап лечения крайне важен, должен быть продуманным и достаточно продолжительным.
Метаболическая терапия при ХЭ направлена на усиление тканевого обмена, активацию энергетических процессов в клетках, устранение последствий гипоксии ткани, в том числе угнетение анаэробного гликолиза. С этой целью используются комплексные метаболические препараты, длительность приема которых должна составлять не менее 3-4 недель. Представителем данной группы препаратов является Актовегин, полученный посредством диализа и ультрафильтрации, в состав которого входят низкомолекулярные пептиды и дериваты нуклеиновых кислот. Актовегин активирует клеточный метаболизм путем увеличения транспорта, накопления и усиления внутриклеточной утилизации глюкозы и кислорода, что приводит к ускорению метаболизма аденозинтрифосфата (АТФ) и повышению энергетических ресурсов клетки. Актовегин удобен для приема. Его выпускают и в таблетках, и в растворе для инъекций. Поэтому женщины, которые по каким-либо причинам не могут лечь в стационар или систематически приезжать на прием, принимают его дома по схеме, расписанной врачом. Актовегин таблетки принимают, не разжевывая, перед едой, запивая небольшим количеством жидкости, по 1 таблетке 2 раза в день в течение 30 дней. При введении в форме инфузии к 200-300 мл основного раствора (0,9% раствор хлорида натрия или 5% раствор декстрозы) добавляют 10-20 мл Актовегина, раствор используют по следующей схеме: внутривенно через день в течение не менее двух недель. Скорость введения: около 2 мл/мин.
Положительное влияние Актовегина на кровообращение связано с улучшением доставки и утилизации кислорода, увеличением перфузии глюкозы и восстановлением аэробного метаболизма как в периферических тканях, так и в нейрональных структурах. Путем активации фермента супероксиддисмутазы препарат оказывает выраженное антиоксидантное и противоишемическое действие. Актовегин хорошо сочетается с другими лекарственными средствами, что очень удобно для комплексного лечения хронического эндометрита.
Традиционно в комплексной терапии ХЭ применяются физические факторы. Наиболее часто в комплексной терапии ХЭ применяют электроимпульсную терапию, интерференционные токи, переменное магнитное поле низкой частоты, ток надтональной частоты. Задачи физиотерапии при ХЭ заключаются в улучшении гемодинамики органов малого таза, стимуляции функции рецепторов, ускорении процессов регенерации эндометрия, повышении иммунологических свойств.
Дискуссионными остаются вопросы о целесообразности использования в комплексном лечении ХЭ циклической гормональной терапии. В настоящее время считается, что при хроническом эндометрите оправдано только дифференцированное назначение гормонотерапии при наличии гипофункции яичников или ановуляции. Потенциал к патологической регенерации ткани, который таит в себе хроническое воспаление, при назначении циклической заместительной гормональной терапии может быть усилен и реализован.
Контроль эффективности терапевтических мероприятий должен проводиться не ранее чем через 2 месяца после окончания лечения с учетом эффекта последействия физиотерапии. При этом оценивается динамика клинических симптомов, проводится ультразвуковое исследование матки (допплерометрия сосудов матки) и аспирационная биопсия эндометрия (на 7-10 день менструального цикла) с последующим морфологическим исследованием эндометрия.
Выбор метода гормональной терапии
Пациенткам, имеющим нормальную толщину эндометрия (более 8 мм) после овуляции и нормальный кровоток в сосудах эндометрия ИР в базальных артериях не более 0,48, в спиральных артериях 0,38, ИР в аркуатных артериях 0,64, ИР в маточной артерии 0,8) при проведении допплерометрии на 20-22 день менструального цикла, в качестве прегравидарной подготовки назначался дидрогестерон внутрь 10 мг 2 раза в сутки с 16-го по 25-й день менструального цикла).
У пациенток со сниженной толщиной эндометрия (менее 6-8 мм) после овуляции препаратами выбора явились Фемостон 2/10 с 1 дня менструального цикла в течение 28 дней по 1 таблетке 1 раз в сутки с добавлением дидрогестерона внутрь 10 мг 1 раз в сутки с 16-го по 25-й день менструального цикла либо фазовая гормонотерапия препаратами: эстрадиол внутрь 2 мг 1 раз в сутки с 1-го по 15-й день менструального цикла и дидрогестерон внутрь 10 мг 2 раза в сутки с 16-го по 25-й день менструального цикла.
Пациенткам, имеющим сочетание снижения толщины эндометрия со снижением кровотока в сосудах эндометрия на фоне заместительной гормональной терапии, назначался курс иглорефлексотерапии, с подбором точек акупунктуры, направленных на улучшение кровотока в матке, а также комплекс метаболической терапии.
Пациенткам группы сравнения на этапе прегравидарной подготовки назначались оральные контрацептивы внутрь по 1 таблетке 1 раз в день с 1-го дня менструального цикла в течение 21-го дня.
Метаболическая терапия при ХЭ направлена на усиление тканевого обмена, активацию энергетических процессов в клетках, устранение последствий гипоксии ткани, в том числе перевод с анаэробного гликолиза на аэробный.
В качестве метаболической терапии применялась внутривенная инфузия аскорбиновой кислоты 5% — 5,0 мл с Рибоксином 2% — 10,0 мл на глюкозе 5% — 200 м в течение 10 дней. Депротеинизированный гемодериват крови телят (Актовегин) 5,0 мл (200 мг) внутривенно капельно на 200 мл физиологического раствора через день 5 процедур. Затем в качестве второго этапа назначался Актовегин в таблетках (200 мг) 3 раза в сутки в протоколе ЭКО до получения результата. Наш опыт показал, что применение препарата Актовегин у пациенток, страдающих ХЭ, за счет метаболического действия, а именно увеличения транспорта и утилизации глюкозы и кислорода клетками тканей, позволяет повысить результативность программ ЭКО.
В терапии хронического эндометрита современная гинекология применяет комплексный подход, включающий противомикробное, иммуномодулирующее, общеукрепляющее, физиотерапевтическое лечение.
Лечение проводится поэтапно. Первым шагом является устранение инфекционных агентов, затем следует курс, направленный на восстановление эндометрия. Обычно применяются антибиотики широкого спектра действия (спарфлоксацин, доксициклин и др.). Восстановительный курс построен на сочетании гормональной (дивигель плюс утрожестан) и метаболической терапии (актовегин, рибоксин, аскорбиновая кислота, витамин Е, вобэнзим).
Купирование маточных кровотечений проводится назначением гормонов или раствора аминокапроновой кислоты (внутривенно или внутриматочно).
Осложнения и профилактика эндометрита
Отдаленными последствиями перенесенного эндометрита служат спаечные процессы внутри матки (внутриматочные синехии), склерозирование полости матки, нарушенное течение менструального цикла, полипы и кисты эндометрия.
Группа была разделена на две подгруппы, одна из которых была взята в программу после проведенной терапии, вторая без проведенного лечения. В первой группе( n10) каждая женщина в течение 2-3 циклов получила антибактериальное лечение, с учетом бакпосева и высеянной микрофлоры; физиолечение, включающее чередование электрофореза и лазеротерапии в первую фазу менструального цикла; циклическую гормональную терапию, которая продолжалась в течение 2-3 менструальных циклов и включала применение прогинова 2-4мг с 1 по 15 день менструального цикла, утрожестан 200 мг с 16 по 25 дмц. Все пациентки получали антиоксидантную терапию (аскорбиновая кислота по 5 драже в день с 1 по 15 день менструального цикла, витамин Е по 5 капсул в день с 15 по 25 дмц), свечи дистрептаза(по схеме 3 дня по 1 свече 3 раза в день p/r,3 дня по 1 св 2 раза в день,3 дня по 1 свече 1 раз в день).Всем 20 пациенткам программа ЭКО проводилась по длинному протоколу. Число аспирированных ооцитов в обеих группах не имело значительных различий и составило от 8 до 15 (в среднем 10,2±1,3). Частота проведенных ИКСИ в обеих группах составляла соответственно 38% в первой и 43,6% во второй группе. Показаниями к проведению ИКСИ явились средняя и тяжелая формы нарушений сперматогенеза. Перенос производился на 3 день культивирования. В первой группе беременность наступила у 3 женщин (30%)и у всех завершилась рождением доношенных детей. Во второй группе у 2 пациенток, составив 20%. У одной женщины беременность остановилась в развитии при сроке сроке 8-9 недель, у второй- завершилась родами в срок.
Таким образом, проведенный анализ продемонстрировал повышение эффективности программ ВРТ и снижение риска невынашивания беременности у женщин с хроническими эндометритами после предварительного комплексного патогенетического лечения.бактероиды снижены что это значит — 8 рекомендаций на Babyblog.ru
Девочки читайте,чтобы быть вкурсе анализа вашего малыша
Как “читать” анализ на дисбактериоз
Копанев Ю.А. Соколов А.Л.
В бланке каждого анализа на дисбактериоз имеются показатели микрофлоры, которые мы расшифруем.
Патогенные энтеробактерии. Обычно этот показатель в бланке анализа идет первым. К этой группе микроорганизмов относятся такие бактерии, которые вызывают острую кишечную инфекцию (сальмонеллы, шигеллы – возбудители дизентерии, возбудители брюшного тифа). Выявление этих микроорганизмов является уже не показателем дисбактериоза, а показателем серьезного инфекционного заболевания кишечника.
Бифидобактерии. Это основные представители нормальной кишечной микрофлоры, количество которых в кишечнике должно быть 95 – 99%. Бифидобактерии выполняют важную работу по расщеплению, перевариванию и всасыванию различных компонентов пищи, например, углеводов; они сами синтезируют витамины, а также способствуют усвоению их из пищи; при участии бифидобактерий происходит всасывание в кишечнике железа, кальция и других важных микроэлементов; бифидобактерии стимулируют моторику кишечной стенки и способствуют нормальному опорожнению кишечника; бифидобактерии нейтрализуют разлчные токсические вещества, попадающие в кишечник извне или образовавшиеся в результате жизнедеятельности гнилостных микроорганизмов. В бланке анализа указывается титр бифидобактерий, который должен быть не меньше 107 – 109. Существенное снижение количества бифидобактерий – всегда признак выраженного дисбактериоза.
Лактобактерии (лактобациллы, молочнокислые микробы, молочнокислые стрептококки). Второй по представительству (5% в совокупности микроорганизмов кишечника) и по значимости представитель нормальной флоры. Лактобактерии или молочнокислые микробы, как показывает их название, вырабатывают молочную кислоту – важнейший компонент для нормальной работы кишечника. Лактобактерии обеспечивают противоаллергическую защиту, способствуют нормальному опорожнению кишечника, вырабатывают высокоактивную лактазу – фермент, расщепляющий молочный сахар (лактозу). В анализе их количество должно быть не меньше 106 – 107. Дефицит лактобактерий может привести к развитию аллергических заболеваний, запорам, лактазной недостаточности.
Кишечная палочка с нормальной ферментативной активностью (эшерихии). Третий представитель нормальной флоры. Количество кишечной палочки среди других бактерий не превышает 1%, но роль ее очень важна, поскольку этот микроб: во-первых, является главным конкурентом условно-патогенной флоры, препятствуя заселению чужеродными микробами кишечной стенки; во-вторых, кишечная палочка забирает из просвета кишечника кислород, который является ядом для бифидобактерий и лактобактерий, таким образом, создаются комфортные условия для основных бактерий кишечной флоры. Следует отметить, что до 6 – 8 месяцев, роль кишечной палочки невелика, а количество ее может колебаться от 100 млн/г до 2 – 3 млрд/г. Ближе к году и в более старшем возрасте общее количество кишечной палочки должно составлять не менее 300 – 400 млн/г (107 – 108). Снижение общего количества кишечной палочки может быть признаком присутствия в кишечнике небактериальных паразитов (глистов, простейших, которые также нуждаются в кислороде, обделяя им кишечную палочку).
Следует отметить, что бактерии нормальной кишечной флоры живут, прикрепляясь к кишечной стенке и образуя пленку, покрывающую кишечник изнутри. Через эту пленку происходит все всасывание в кишечнике. Бактерии нормальной кишечной микрофлоры обеспечивают по совокупности 50 – 80% всего пищеварения, а также выполняют защитные (в том числе противоаллергические) функции, нейтрализуют действие чужеродных и гнилостных бактерий, способствуют опорожнению кишечника, обеспечивают адаптацию к питанию и к внешним воздействиям.
Кишечная палочка со сниженной ферментативной активностью. Это неполноценная кишечная палочка, которая не представляет никакого вреда, но при этом не выполняет своих полезных функций. Присутствие данного показателя в анализе является признаком начинающегося дисбактериоза, а также как и снижение общего количества кишечной палочки может быть косвенным признаком присутствия в кишечнике глистов или простейших.
В некоторых анализах описываются бактероиды, роль которых неясна, но известно, что это – не вредные бактерии, обычно их количество не имеет практического значения.
Все остальные показатели микрофлоры являются условно-патогенной флорой. В самом термине “условно-патогенный” обозначена суть данных микробов. Они становятся патогенными (нарушающими нормальные функции кишечника) при определенных условиях: повышение их абсолютного количества или процентного соотношения к нормальной флоре, при неэффективности защитных механизмов или снижении функции иммунной системы. Условно-патогенная флора – это лактозонегативные энтеробактерии (клебсиелла, протей, цитробактеры, энтеробактеры, гафнии, серрации), гемолизирующая кишечная палочка и различные кокки (энтерококки, эпидермальные или сапрофитные стафилококки, золотистый стафилококк). Кроме того, к условно-патогенным относятся клостридии, которые высеваются не во всех лабораториях. Условно-патогенная флора внедряется, конкурируя с полезными бактериями, в микрбную пленку кишечника, заселяет кишечную стенку и вызывает нарушения работы всего желудочно-кишечного тракта. Дисбактериоз кишечника с повышенным содержанием условно-патогенной флоры может сопровождаться аллергическими кожными реакциями, нарушениями стула (запоры, поносы, зелень и слизь в кале), болями в животе, вздутиями живота, срыгиваниями, рвотами. При этом обычно температура тела не повышается.
Кокковые формы в общей сумме микробов. Самыми безобидными представителями условно-патогенной флоры являются энтерококки. Они наиболее часто встречаются в кишечнике у здоровых людей, их количество до 25% не представляет угрозы здоровью. Если количество превышает 25% (более 107), это чаще всего связано со снижением нормальной флоры. В редких случаях повышение количества энтерококков является главной причиной дисфункции, связанной с дисбактериозом.
Эпидермальный (или сапрофитный) стафилококк (S. epidermidis, S. saprophyticus). Эти виды стафилококков могут вызывать нарушения, но их количество до 25% является допустимым.
Процент гемолизирующих кокков по отношению ко всем кокковым формам. Даже среди относительно безобидных кокков, названных выше, могут встречаться более патогенные, что и обозначается в этой позиции. Если общее количество кокков составляет, например, 16%, а процент гемолизирующих кокков – 50%, это значит, что половина из 16% — более вредоносные кокки, а их процент по отношению к нормальной флоре составляет 8%.
Золотистый стафилококк (S. aureus). Один из самых неприятных (наряду с гемолизирующей кишечной палочкой, протеем и клебсиеллой) представителей условно-патогенной флоры. Даже небольшие его количества могут вызвать выраженные клинические проявления, особенно у детей первых месяцев жизни. Поэтому обычно в нормах, приведенных в бланке анализа, указывается, что его быть не должно (на самом деле допустимы количества, не превышающие 103). Патогенность золотистого стафилококка напрямую зависит от состояния нормальной флоры: чем больше бифидобактерий, лактобактерий и нормальной кишечной палочки, тем меньше вреда от стафилококка. Присутствие его в кишечнике может приводить к аллергическим реакциям, гнойничковым кожным высыпаниям, дисфункции кишечника. Стафилококки являются распространенными микробами окружающей среды, в частности, они в больших количествах живут на коже и слизистых оболочках верхних дыхательных путей. Ребенку они могут попадать через грудное молоко. Наиболее подвержены заражению стафилококками ослабленные дети (проблемная беременность, недоношенность, кесарево сечение, искусственное вскармливание, применение антибиотиков – факторы риска ослабления функций иммунной системы). Важно понимать, что стафилококки как и другие условно-патогенные бактерии проявляют себя при определенных условиях, главным из которых является ослабление иммунитета, поэтому важным при лечении дисбактериоза, ассоциированного со стафилококком, является проведение иммунокоригирующей терапии.
Гемолизирующая кишечная палочка. Является представителем лактозонегативных энтеробактерий, но выделяется отдельно в связи с распространенностью и значимостью. В норме должна отсутствовать. К данному микробу применимо практически все, сказанное про золотистый стафилококк. То есть, она может вызывать аллергические и кишечные проблемы, очень распространена в окружающей среде (правда, практически никогда не встречается в грудном молоке), вызывает проблемы у ослабленных детей, требует иммунокоррекции. Следует отметить, что термин “гемолизирующая” не означает, что имеется какое-то влияние на кровь. Условно-патогенная флора при дисбактериозе не должна преодолевать кишечную стенку и попадать в кровь. Это возможно только при крайне выраженных формах дисбактериоза у детей с выраженными иммунодефицитами, как правило, представляющих угрозу для жизни. К счастью, такие состояния встречаются нечасто.
Лактозонегативные энтеробактерии. Большая группа условно-патогенных бактерий большей или меньшей степени патогенности. Их количество не должно превышать 5% (или в титрах: 103 – 106 – умеренное повышение, более 106 – существенное повышение). Наиболее неприятными бактериями из этой группы являются протеи (чаще всего с ними связаны запоры) и клебсиеллы (являются прямыми антагонистами (конкурентами) лактобактерий, что приводит к развитию аллергии и запоров, а также к проявлениям лактазной недостаточности). Часто в бланке анализа указывается общее количество лактозонегативных энтеробактерий (наиболее информативно процентное соотношение), а затем идет расшифровка:
Клебсиеллы;
Протеи;
Хафнии;
Серрации;
Энтеробактеры;
Цитробакеры.
Обычно какие-то количества этих бактерий постоянно живут в кишечнике, не вызывая проблем. В нормах могут быть указаны цифры от 103 до 106, являющиеся допустимыми.
Грибы рода Candida. Допустимо присутствие до 104. Повышение этого параметра может быть после применения антибиотиков. Если количество грибов повышено, а количество нормальной кишечной флоры резко снижено, при этом отмечается кандидоз (молочница) видимых слизистых оболочек (ротовая полость, половые органы) – это проявления системного кандидоза, то есть имеется инфицирование грибами кишечника. Если же количество грибов в анализе на дисбактериоз увеличено, но нет снижения нормальной кишечной флоры, это свидетельствует о том, что грибы живут на коже вокруг заднего прохода, а не в кишечнике, в этом случае достаточно наружной терапии с использованием противогрибковых мазей или кремов.
Клостридии. Из-за технических сложностей и небольшого практического значения определяются не во всех лабораториях. Допустимое количество до 107. Проявляют патогенность обычно в комплексе с другой условно-патогенной флорой, редко изолированно вызывают проблемы (чаще всего – разжижение стула, понос). Их количество зависит от функции местного иммунитета кишечника.
Прочие микроорганизмы. В данном параметре описываются редко встречающиеся виды бактерий, самым опасным из которых является синегнойная палочка (Pseudomonas aerugenosa). Чаще всего, микроорганизмы, описанные в этой позиции анализа не имеют практического значения.
Термин “abs” обозначает отсутствие данного микроорганизма, также употребляется “не обнаружено”.
Дисбактериоз кишечника — Клиника Здоровье 365 г. Екатеринбург
Микробиоцеоз – это сообщество микроорганизмов, обитающих на территориально ограниченном участке с однородными условиями жизни. Бактерии нормальной кишечной флоры живут, прикрепляясь к кишечной стенке и образуя пленку, покрывающую кишечник изнутри. Через эту пленку происходит все всасывание в кишечнике. Бактерии нормальной кишечной микрофлоры обеспечивают по совокупности 50 – 80% всего пищеварения, а также выполняют защитные функции, нейтрализуют действие чужеродных и гнилостных бактерий. Бифидобактерии. Это основные представители нормальной кишечной микрофлоры, количество которых в кишечнике должно быть 95 – 99%. Бифидобактерии выполняют важную работу по расщеплению, перевариванию и всасыванию различных компонентов пищи, например, углеводов; они сами синтезируют витамины, а также способствуют усвоению их из пищи. При участии бифидобактерий происходит всасывание в кишечнике железа, кальция и других важных микроэлементов; нейтрализуют различные токсические вещества. В бланке анализа указывается титр бифидобактерий, который должен быть не меньше 107 – 108. Существенное снижение количества бифидобактерий – признак выраженного дисбактериоза.
Лактобактерии (лактобациллы, молочнокислые микробы, молочнокислые стрептококки).
Второй по представительству (5% в совокупности микроорганизмов кишечника) и по значимости представитель нормальной флоры. Лактобактерии или молочнокислые микробы вырабатывают молочную кислоту – важнейший компонент для нормальной работы кишечника. В результатах исследования их количество должно быть не меньше 106 – 107. Дефицит лактобактерий может привести к развитию аллергических заболеваний, запоров, лактазной недостаточности.
Кишечная палочка с нормальной ферментативной активностью (эшерихии).
Третий представитель нормальной флоры. Количество кишечной палочки среди других бактерий не превышает 1%, но роль ее очень важна. Во-первых, кишечная палочка является главным конкурентом условно-патогенной флоры, препятствуя заселению чужеродными микробами кишечной стенки; во-вторых, кишечная палочка забирает из просвета кишечника кислород, который является ядом для бифидобактерий и лактобактерий. Снижение общего количества кишечной палочки может быть признаком присутствия в кишечнике небактериальных паразитов (глистов, простейших, которые также нуждаются в кислороде, обделяя им кишечную палочку). Кишечная палочка со сниженной ферментативной активностью. Это неполноценная кишечная палочка, которая не представляет никакого вреда, но при этом не выполняет своих полезных функций. Присутствие данного показателя в анализе является признаком начинающегося дисбактериоза, а также как и снижение общего количества может быть косвенным признаком присутствия в кишечнике глистов или простейших.
В некоторых анализах описываются бактероиды, роль которых неясна, но известно, что это – не вредные бактерии, обычно их количество не имеет практического значения. Все остальные показатели микрофлоры являются условно-патогенной флорой. Они становятся патогенными (нарушающими нормальные функции кишечника) при определенных условиях: повышение их абсолютного количества или снижении функции иммунной системы.
Условно-патогенная флора – это лактозонегативные энтеробактерии (клебсиелла, протей, цитробактеры, энтеробактеры, гафнии, серрации), гемолизирующая кишечная палочка и различные кокки (энтерококки, эпидермальные или сапрофитные стафилококки, золотистый стафилококк). Кроме того, к условно-патогенным относятся клостридии, которые высеваются не во всех лабораториях. Условно-патогенная флора внедряется, конкурируя с полезными бактериями, в микробную пленку кишечника, заселяет кишечную стенку и вызывает нарушения работы всего желудочно-кишечного тракта. Дисбактериоз кишечника с повышенным содержанием условно-патогенной флоры может сопровождаться аллергическими кожными реакциями, нарушениями стула (запоры, поносы, зелень и слизь в кале), болями в животе, вздутиями живота, срыгиваниями, рвотами.
Кокковые формы в общей сумме микробов.
Самыми безобидными представителями условно-патогенной флоры являются энтерококки. Они наиболее часто встречаются в кишечнике у здоровых людей, их количество до 106 — 107 не представляет угрозы здоровью. Если количество превышает 25% (более 107), это чаще всего связано со снижением нормальной флоры. Эпидермальный (или сапрофитный) стафилококк (S. epidermidis, S. saprophyticus). Эти виды стафилококков могут вызывать нарушения, но их количество до 104 является допустимым. Золотистый стафилококк (S. aureus). Один из самых неприятных (наряду с гемолизирующей кишечной палочкой, протеем и клебсиеллой) представителей условно-патогенной флоры. Даже небольшие его количества могут вызвать выраженные клинические проявления, особенно у детей первых месяцев жизни. Поэтому обычно в нормах, приведенных в бланке анализа, указывается, что его быть не должно (на самом деле допустимы количества, не превышающие 103). Патогенность золотистого стафилококка напрямую зависит от состояния нормальной флоры: чем больше бифидобактерий, лактобактерий и нормальной кишечной палочки, тем меньше вреда от стафилококка. Гемолизирующая кишечная палочка. Является представителем лактозонегативных энтеробактерий, но выделяется отдельно в связи с распространенностью и значимостью. В норме должна отсутствовать. К данному микробу применимо практически все, сказанное про золотистый стафилококк. То есть, она может вызывать аллергические и кишечные проблемы, очень распространена в окружающей среде (правда, практически никогда не встречается в грудном молоке), вызывает проблемы у ослабленных детей, требует иммунокоррекции. Следует отметить, что термин “гемолизирующая” не означает, что имеется какое-то влияние на кровь. Лактозонегативные энтеробактерии. Большая группа условно-патогенных бактерий большей или меньшей степени патогенности. Их количество не должно превышать (в титрах: 103 – 106 ). Наиболее неприятными бактериями из этой группы являются протей (чаще всего с ними связаны запоры) и клебсиеллы (являются прямыми антагонистами (конкурентами) лактобактерий, что приводит к развитию аллергии и запоров, а также к проявлениям лактазной недостаточности). Часто в бланке анализа указывается общее количество лактозонегативных энтеробактерий (наиболее информативно процентное соотношение), а затем идет расшифровка: клебсиеллы, протей, хафнии, серрации, энтеробактерии и цитробактерии. Обычно какие-то количества этих бактерий постоянно живут в кишечнике, не вызывая проблем. В нормах могут быть указаны цифры от 104 до 105, являющиеся допустимыми.
Грибы рода Candida.
Допустимо присутствие до 103. Повышение этого параметра может быть после применения антибиотиков. Если количество грибов повышено, а количество нормальной кишечной флоры резко снижено, при этом отмечается кандидоз (молочница) видимых слизистых оболочек (ротовая полость, половые органы) – это проявления системного кандидоза, то есть имеется инфицирование грибами кишечника. Если же количество грибов в анализе на дисбактериоз увеличено, но нет снижения нормальной кишечной флоры, это свидетельствует о том, что грибы живут на коже вокруг заднего прохода, а не в кишечнике, в этом случае достаточно наружной терапии с использованием противогрибковых мазей или кремов.
Клостридии. Допустимое количество до 105. Проявляют патогенность обычно в комплексе с другой условно-патогенной флорой, редко изолированно вызывают проблемы (чаще всего – разжижение стула, понос). Их количество зависит от функции местного иммунитета кишечника.
Прочие микроорганизмы.
В данном параметре описываются редко встречающиеся виды бактерий, самым опасным из которых является синегнойная палочка (Pseudomonas aerugenosa). Чаще всего, микроорганизмы, описанные в этой позиции, не имеют практического значения. Термин “abs” обозначает отсутствие данного микроорганизма, также употребляется “не обнаружено”.
➤ Микробиоценоз влагалища и факторы, влияющие на его состояние
1. Анкирская А.С., Муравьева В.В. Интегральная оценка состояния микробиоты влагалища, диагностика оппортунистических вагинитов. Медицинская технология. М.: ФГБУ «НЦ АГиП им. В.И. Кулакова» Минздравсоцразвития России, 2011.
2. Коршунов В.М., Володин Н.Н., Ефимов Б.А. и др. Микроэкология влагалища. Коррекция микрофлоры при вагинальных дисбактериозах: Учебное пособие. М.: ВУНМЦ МЗ РФ, 1999. 80 с.
3. Титова С.И., Гончарова Н.Г. Оценка клинической эффективности, безопасности и переносимости комбинации пребиотика и энтеросорбента в терапии бактериального вагиноза // Лечащий врач. 2008. №10.
4. Каминский В.В., Однокоз Т.А. и соавт. Сборник клинических исследований препаратов Лактофильтрум и Фильтрум. М., 2008.
5. Ворошилина Е.В., Тумбинская Л.В., Донников А.Е., Плотко Е.Э., Хаютин Л.В. Биоценоз влагалища с точки зрения количественной ПЦР: изменения и коррекция во время беременности // Инфекции в гинекологии. 2010. Т. 68. №3. С. 108–111.
6. Кира Е.Ф. Бактериальный вагиноз. М.: ООО «Нева-Люкс», 2001. 364 c.
7. Липова Е.В., Болдырева М.Н., Трофимов Д.Ю., Витвицкая Ю.Г. Урогенитальные инфекции, обусловленные условно-патогенной биотой у женщин репродуктивного возраста (клинико-лабораторная диагностика) // Пособие для врачей. М., 2009. С. 30.
8. Плотко Е.Э., Донников А.Е., Ворошилина Е.С., Хаютин Л.В., Тумбинская Л.В. Биоценоз влагалища с точки зрения количественной ПЦР: что есть норма? // Акушерство и гинекология. 2011. №1. С. 66–70.
9. Прилепская В.Н., Байрамова Г.Р. Вульвовагинальный кандидоз. Клиника, диагностика, принципы терапии. М.: ООО «ГЭОТАР-Медиа», 2010. 80 c.
10. Радзинский В.Е., Ордиянц И.М. Профилактика послеродовых инфекций у женщин с бактериальным вагинозом // Гинекология. 2006. Т.8. №1. С. 14–16.
8. Сидорова И.С. Микробиоценоз половых путей женщин репродуктивного возраста // Акушерство и гинекология. 2005. №2. С. 7–9.
9. Malazy O.T., Shariat M., Heshmat R., Majlesi F. et al. Vulvovaginal candidiasis and its related factors in diabetic women // Taiwan J Obstet Gynecol. 2007, 46(4): 399–404.
10. Anderson B.L., Cu-Uvin S., Raker C.A. et al. Subtle perturbations of genital microflora alter mucosal immunity among low-risk pregnant women // Acta Obstet. Gynecol. Scand. 2011. Vol. 90, №5. P. 510.
11. Blum S., Schiffrin E.J. Intestinal microflora and homeostasis of the mucosal immune response: implications for probiotic bacteria? // Current issues in intestinal microbiology. 2003. Vol. 4, №2. P. 53–60.
12. Bradshaw C.S., Tabrizi S.N., Fairley C.K. et al. The association of Atopobium vaginae and Gardnerella vaginalis with bacterial vaginosis and recurrence after oral metronidazole therapy // The Journal of infectious diseases. 2006. Vol. 194, №6. P. 828–836.
13. Donders G.G., Bellen G., Mendling W. Management of recurrent vulvo-vaginal candidosis as a chronic illness // Gynecologic and obstetric investigation. 2010. Vol. 70, №4. P. 306–321.
14. Krauss-Silva L., Moreira M.E., Alves M.B. et al. Randomized controlled trial of probiotics for the prevention of spontaneous preterm delivery associated with intrauterine infection: study protocol // Reproductive health. 2010. Vol. 7. P. 14.
15. Levison M.E., Corman L.C., Carrington E.R., Kaye D. Quantitative microflora of the vagina // American journal of obstetrics and gynecology. 1977. Vol.127, №1. P. 80–85.
16. Linhares I.M., Giraldo P.C., Baracat E.C. New findings about vaginal bacterial flora // Revista da Associacao Medica Brasileira. 2010. Vol.56, №3. P. 370–374.
17. Nugent R.P., Krohn M.A., Hillier S.L. Reliability of diagnosing bacterial vaginosis is improved by a standardized method of gram stain interpretation. // Journal of clinical microbiology.-1991.- Vol.29, № 2.- P. 297-301.
18. Orendi J.M., Coetzee N., Ellington M. J. et al. Community and nosocomial transmission of Panton-Valentine leucocidin-positive community-associated meticillin-resistant Staphylococcus aureus: implications for healthcare. // The Journal of hospital infection.-2010.- Vol.75, № 4.- P. 258-264.
19. Roberts C.L., Morris J.M., Rickard K.R. et al. Protocol for a randomised controlled trial of treatment of asymptomatic candidiasis for the prevention of preterm birth. // BMC pregnancy and childbirth.-2011.- Vol. 11.- P. 19.
20. Sherrard J., Donders G., White D., Jensen J.S. European (IUSTI/WHO) guideline on the management of vaginal discharge, 2011. // International journal of STD & AIDS.-2011.- Vol.22, № 8.- P. 421-429.
21. Verhelst R., Verstraelen H., Claeys G. et al. Cloning of 16S rRNA genes amplified from normal and disturbed vaginal microflora suggests a strong association between Atopobium vaginae, Gardnerella vaginalis and bacterial vaginosis. // BMC microbiology.-2004.- Vol.4.- P. 16.
22. Zodzika J., Rezeberga D., Jermakova I. et al. Factors related to elevated vaginal pH in the first trimester of pregnancy. // Acta obstetricia et gynecologica Scandinavica. 2011. Vol. 90, №1. P. 41–46.
Микробиом кишечника | Лабораторія Др. Рьодгера
Соотношение Firmicutes/BacteroidetesКорреляция между соотношением Firmicutes / Bacteroidetes и массой тела человека была обнаружена во многих исследованиях в последние годы: чем меньше доля Firmicutes, то есть чем больше доля Bacteroidetes, тем меньше масса тела.
На рис. 6 показан пример американского исследования, в котором доля Bacteroidetes увеличивается с увеличением потери веса, как при диете со снижением жиров, так и при диете со снижением углеводов(Ley R. et al., 2006).
Количество короткоцепочечных жирных кислот в кишечнике — это подход для объяснения высокой доли Firmicutes у людей с избыточным весом. В исследовании на животных было обнаружено значительно большую концентрацию короткоцепочечных жирных кислот (KKFS) в кале мышей с ожирением. Энергия от KKFS может составлять 8-10% от общего количества и, таким образом, отражать более высокую генерацию энергии из неусвояемых углеводов, которые образуются бактериями Firmicutes(Duncan S. et al., 2007).
Вредные пищевые привычки, которые негативно влияют на развитие микробиома, уже могут проявляться в грудном возрасте — из-за употребления сбалансированной пищевой смеси в первые месяцы жизни. В подростковом и взрослом возрасте на развитие микробиома негативно влияют: стрессы, продукты и ингредиенты промышленного производства, такие как подсластители, алкоголь или токсины окружающей среды в продуктах. Кроме того, нерациональная терапия антибиотиками, не разграничивает патогенные и физиологические микробы, часто является основной причиной развития дисбактериоза. Во время и после приема антибиотиков можно применять терапию пробиотиками или синбиотики для предотвращения дисбактериоза или восстановления «здорового» микробиома. (См. Раздел «Терапия микробиома»).
Риски заболеваний, связанные с микробиомомНарушение микробиома (дисбактериоз), уменьшение разнообразия или неправильную колонизацию можно считать факторами риска многочисленных заболеваний. К ним относятся: нарушенная перистальтика (диарея или запор), синдром раздраженного кишечника, метаболические заболевания, такие как сахарный диабет, ожирение или нарушение жирового обмена, аутоиммунные заболевания, такие как аллергия или псориаз, хронические воспалительные заболевания кишечника(Colitis ulcerosa или болезнь Крона), злокачественные или даже неврологично- психические заболевания (Frank D. и др., 2011). Основа для этих процессов может быть различного типа. Возможно взаимодействие между рецепторами эпителиальных и иммунных клеток слизистой оболочки кишечника и продуктами бактериального метаболизма, такими как короткоцепочечные жирные кислоты или липополисахарида.
Перечень заболеваний, связанных с нарушением микробиома:■ Метаболические заболевания
Ожирение, диабет второго типа, сердечно-сосудистые заболевания, безалкогольная жировая болезнь печени, алкогольная жировая болезнь печень
■ Синдром раздраженного кишечника
Раздраженный кишечник, Leaky-Gut-синдром, непереносимость гистамина, непереносимость пищи, SIBOS
■ Воспалительные заболевания кишечника
Хронические воспалительные заболевания кишечника, колоректальный рак, дисбактериоз, сниженная резистентность к колонизации, восприимчивость желудочно-кишечного тракта к инфекциям
■ Аутоиммунные заболевания
Целиакия, ревматоидный артрит, псориаз, аллергия / астма, сахарный диабет 1 типа
■ Неврологические заболевания
Депрессия, синдром хронической усталости, расстройство спектра аутизма, болезнь Паркинсона, болезнь Альцгеймера
Кроме того, можно проводить исследования на потенциально патогенные бактерии. Среди них Clostridium difficile, что образует токсин A / B, который вызывает диарею, вызванную антибиотиками, и Helicobacter pylori, который является причиной язвы желудка и двенадцатиперстной кишки и рака желудка. Особое значение при Helicobacter pylori имеет антибиотикорезистентнисть к кларитромицину, которая становится все более распространенной.
Несмотря на то, что у большинства пациентов ожирение вызывается потреблением высококалорийной пищи и недостаточными физическими нагрузкам, некоторые исследования показали корреляцию между бактериями и весом пациента. Например, было замечено, что низкая колонизация бактероидами и преобладание штамма Firmicutes характерные для больных ожирением (Cani P. 2013).
В другом исследовании, в котором приняли участие 115 пациентов, с помощью qPCR удалось показать, что сниженная доля Bacteroidetes, Bifidobacterium spp. и Methanobrevibacter smithii и увеличенная доля Lactobacillus reuteri ассоциировались с лишним весом (Million M et al. 2012).
John K. Dibaise (Dibaise JK. Et al. 2008), который изучал результаты ранее известных исследований, обнаружил связь между составом микробиома кишечника и развитием факторов риска метаболического синдрома.
Исследования показали, что у мышей традиционно выведенных было на 40% больше содержания жира по сравнению со « стерильными» мышами, хотя они получали меньше пищи. После того, как кишечные микробы обычно разведенных животных были переданы «стерильным» животным, содержание жира у последних увеличился на 60% в течение 2 недель. Состав корма и его количество не менялись.
Увеличение жира также было связано с повышенной резистентностью к инсулину, увеличением адипоцитов, и повышением уровня глюкозы и лептина в крови.
Механизм этих изменений основан на ферментации непереваренных полисахаридов, благодаря чему образуются рассасываюшиеся моносахариды, и короткоцепочечные жирные кислоты, которые поглощаются печенью и липоцитами жировой ткани, превращаются в жир и накапливаются. Увеличение веса также индуцируется угнетением фактора жировой ткани, индуцированного голоданием (FIAF, fasting induced adipocyte Factors). Этот фактор подавляет активность липопротеиновой липазы (ЛПЛ), функцией которой является освобождение жирных кислот из липопротеинов. Подавленное FIAF увеличивает активность ЛПЛ и, таким образом, количество свободных жирных кислот.
Кроме вышеупомянутых механизмов, увеличение веса, вызванное дисбактериозом, и связанная с ним инсулиновая резистентность способствуют хроническому системному воспалению.
Ряд экспериментов на животных показал, что липополисахарида (ЛПС) с клеточной стенки грамотрицательных бактерий, которые считаются эндотоксинами, играют важную роль в развитии хронического воспаления и метаболического синдрома при рационе с высоким содержанием жира. Было замечено, как изменение концентрации ЛПС влияло на состав кишечных бактерий при рационе с высоким содержанием жира. Ендотоксинемия снижала как грамотрицательные Bacteroides spp., так и грамположительные Eubacterium rectale, Clostridium coccoides и Bifdobacteria spp. В целом это привело к увеличению доли грамотрицательных бактерий.
Ендотоксинемия, путем связывания ЛПС с поверхностью CD14-лимфоцитов, стимулирует выработку провоспалительных
цитокинов, таких как фактор некроза опухоли (ФНО), ингибитор активатора плазминогена тип-1 (PAI-1), интерлейкины IL-1 и IL-6. Считается, что эта хроническая метаболическая ендотоксинемия через систему LPS / CD-14 индуцирует чувствительность к инсулину и, следовательно, развитие ожирения и диабета.
В исследованиях больных сахарным диабетом 2 типа было обнаружено более высокий уровень ЛПС в крови, чем у пациентов без диабета. Кроме того, лечение полимиксином В, специальным антибиотиком против грамотрицательных бактерий, показало снижение влияния ЛПС и образования жировой ткани печени.
Быстрый рост ожирения, инсулинорезистентности и сахарного диабета среди детей и подростков особенно наблюдается в индустриально развитых странах. Эти заболевания цивилизации, согласно литературным данным, связанные с дисбактериозом и неправильной колонизацией. Считается, что причиной является использование антибиотиков. Роль пенициллина, хинолонов, цефалоспоринов и макролидов изучалась исследовательской группой Медицинской школы при Университете Пенсильвании. Были проанализированы данные примерно 10 млн пациентов в Великобритании. Результат: риск заболевания увеличивается с количеством антибиотикотерапии; противогрибковая или противовирусная терапия, с другой стороны, не влияют на частоту и прогрессирование этих заболеваний (Boursi B. et al. 2015).
Воспалительные заболевания кишечника
Болезнь Крона или Colitis ulcerosa являются серьезными диагнозами для пациентов. Их типичными симптомами являются диарея, боль в животе и перианальные кровотечения, которые возникают периодически. Хотя первичные проявления имеют место в кишечнике, поражаются также суставы, глаза и кожа. Эти кишечные заболевания относительно распространены. В Европе они встречаются с распространенностью 1: 198 при Colitis ulcerosa и 1: 310 при болезни Крона. Причина — нарушение барьера слизистой оболочки для комменсальних кишечных бактерий (Wehkamp J. et al., 2016). При обоих кишечных заболеваниях наблюдается снижение видового разнообразия, как отражение нарушенной кишечной флоры .
Это нарушение приводит к низкой колонизации бактериями Roseburia spp., Ruminococcus spp., Akkermansia muciniphila, Faecalibacterium prausnitzii и Odoribacter splanchnicus, которые разрушают существующий слой слизи, но и индуцируют быстрое восстановление этого защитного слоя слизи, образуя короткоцепочечные жирные кислоты, такие как масляная кислота. Если жирных кислот не хватает, как важного источника энергии для колоноцитов, то пролиферация, дифференцировка и выработки муцина и дефензины нарушаются и возможна инвазия бактерий. Роды Escherichia и Shigella играют здесь особую роль, поскольку они активируют Toll-подобные рецепторы 4 (TLR4) через липополисахариды и запускают воспалительный процесс (Morgan X. et al. 2012).
Кроме того, предполагается, что одной из возможных причин Colitis ulcerosa является усиленная колонизация Fusobacterium spp. (Yukawa T. et al. 2013)
Синдром раздраженного кишечника
Наиболее частым нарушением у пациентов с проблемами желудочно-кишечного тракта является синдром раздраженного кишечника. Это нарушение функции кишечника, которое поражает женщин примерно вдвое чаще, чем у мужчин. Симптомы раздраженного кишечника включают периодические боли в животе, диарею, запоры и метеоризм. Причинами часто называют нарушение энтеральной моторики, висцеральную гиперчувствительность, семейную предрасположенность и конституционно психологические стрессовые факторы, так как, несмотря на тщательное медицинское обследование, причину, связанную с органами, не было определено.
В базовых исследованиях микробиом человека связь этих жалоб с составом кишечных бактерий наблюдался у многих пациентов. По сравнению с контрольной группой пациентов без симптомов, эти различия приводят к примерно в 1,5 раза увеличение количества бактерий родов Dorea, Ruminococcus и Clostridium spp . а также значительного сокращения родов Bifdobacterium spp. и Faecalibacterium prausnitzii (Rajilic-Stojanovic M.et al. 2013).
Рак толстой кишки
Рак толстой кишки (колоректальная карцинома, colorectale carcinoma = CRC) является вторым по распространенности в Германии типом опухоли. Рак толстой кишки имеет самый высокий уровень заболеваемости в Западной Европе. Кроме генетических факторов (примерно 10% случаев), курения, пищевых ошибок и употребления алкоголя, онкогенезу особенно способствуют длительные хронические воспалительные заболевания кишечника и дисбактериоз.
Многочисленные исследования последних лет показали, что дисбиотические микробиом имеет причинно-следственное значение: отдельные бактерии, такие как Fusobacterium nucleatum или их фактор вирулентности FadA- протеин, могут способствовать развитию рака толстой кишки и его метастазированию (Flanagan L. et al., 2014).
Текущие исследования также показали, что высокая относительная доля Providencia spp . значительно чаще встречается у пациентов с колоректальным раком, чем у здоровых людей. Колонизация кишечника кишечной палочкой (Escherichia coli) типа NC101 или Bacteroides fragilis также может иметь канцерогенное действие. Уменьшение разнообразия и значительно снижена защитная флора слизистой является одним из важнейших факторов риска развития опухоли. Akkermansia muciniphila и Faecalibacterium prausnitzii в настоящее время считаются важнейшим защитным фактором против неоплазии ( Burns M. et al. 2015)
Ревматоидный артрит
Ревматоидный артрит (РА) — одно из самых распространенных хронических воспалительных аутоиммунных заболеваний. В одном исследовании были проанализированы образцы кала пациентов с РА и сравнению со здоровой популяцией. Хотя на уровне штаммов не было значительных различий, четкое отклонения рода Prevotella spp . выявлено у 75% пациентов с РА. Кроме того, Prevotella copri был определен как самый распространенный вид, и было обнаружено два патофизиологические явления: в микробиоме, где преобладают Prevotella — в отличие от микробиома, где преобладают бактероиды — в сыворотке крови наблюдается повышение уровня провоспалительного триметиламина N-Oксида (TMAO). Во-вторых в микробиоме наблюдался пониженный биосинтез фолиевой кислоты (ТHF).
Метотрексат (MTX) — антагонист THF и ингибитор дигидрофолатредуктазы (DHF) — используется для лечения РА. Открытие пониженного биосинтеза фолиевой кислоты в метагеномах, где преобладает Prevotella привело к интересному наблюдения у пациентов с РА, которым проводилось лечение МТХ. Около половины пациентов ответили адекватно на первичную дозу МТХ. Другой половине пациентов часто требовалась дополнительная внутривенная доза МТХ для достижения желаемого эффекта.
Анализ кишечного микробиома показал, что в первой группе пациентов был метагеном, в котором преобладала Prevotella; группа пациентов с повышенной потребностью в MTX, имела метагеном, в котором преобладали бактероиды (Scher J . et al., 2013). В дальнейшем альтернативные варианты терапии могут быть результатом метагеномного исследования кишечной флоры.
Сердечно-сосудистые заболевания
Сердечно-сосудистые заболевания (ССЗ) — самые распространенные заболевания во всем мире. Прогрессированию этих заболеваний способствует провоспалительный триметиламин N-оксид (TMAO). TMAO производится в печени с триметиламина (TMA), который производится бактериями, колонизирующими толстую кишку. В более высоких концентрациях ТМА определяется особенно у людей с высоким потреблением мяса (Ettinger G. et al., 2014).
В исследованиях метаболических свойств кишечных бактерий было замечено, что некоторые бактерии производят ТМА из субстрата холина. Штаммы Firmicutes и Proteobacteria были определены как главные производители TMA (Kymberleigh A. et al. 2015). Поэтому эти бактерии конкурируют с человеком за субстрат холина, из которого в организме вырабатывается важный нейромедиатор ацетилхолин. Уменьшенное потребление холина приводит к сбоям в работе различных органов и способствует развитию ССЗ, раку или повреждению паренхимы печени.
При сбалансированном питании дополнительного потребления холина не требуется, если в рационе содержится достаточное количество метионина и фолиевой кислоты. Кроме того, знание возможных сердечно-сосудистых рисков со стороны кишечной флоры могут помочь предотвратить прогрессирование этих заболеваний путем целенаправленной терапии пробиотиками или пребиотиками.
Кальций-оксалатные мочевые камни
Щавелевая кислота — малорастворима, так что она легко образует кристаллы в моче. В сочетании с кальцием образуются камни оксалата кальция, что может привести, в случае выраженного нефролитиаза, к болезненному застою мочи, воспалению мочевого пузыря и/или почечной лоханки, к пиелонефриту или постренальной почечной недостаточности. Это образование почечных камней поражает примерно 10% населения Северной Америки и Европы.
Oxalobacter formigenes, грамотрицательный и анаэробный микроб, приводит к деградации оксалатов в формиат и СО2 в кишечнике, поэтому считается нефропротективный микробом. Дети в возрасте от 6 до 8 лет почти полностью колонизированы этим микробом. На поздней фазе жизни люди теряют этот микроб, поэтому его присутствие в кишечнике оценивается примерно на 70%. Самой распространенной причиной потери этого микроорганизма в кишечнике есть часто ненужное широкое применение антибиотиков (Stewart C. et al. 2004).
Здоровый образ жизни и уменьшение часто ненужного потребления антибиотиков при инфекционных заболеваниях вирусного происхождения могут дать возможность спонтанной реколонизации флоры кишечника этим микробом.
Нейропсихические расстройства
В последние годы было проведено исследование влияния кишечной флоры на развитие таких заболеваний, как аутизм и деменция, а также нервно-психические расстройства. Было замечено, что воспалительные процессы кишечника способствуют развитию шизофрении, депрессии и биполярных расстройств. Эти заболевания в сочетании с изменениями кишечной флоры могут свидетельствовать, что изменения в желудочно-кишечном тракте играют важную роль при нервно-психических заболеваниях.
При исследовании образцов кала у детей-аутистов по сравнению со здоровой группой, определялось почти 10-кратное повышение относительной частоты рода Clostridium spp. Кроме того, произошли изменения распределения частоты штаммов Bacteroidetes и Firmicutes в пользу Bacteroidetes и других кишечных бактерий, таких как Bifidobacterium, Lactobacillus, Suterrella, Prevotella, Ruminococcus и Alcaligenaceae.Кроме того, наблюдались хронические кишечные инфекции, вызванные Clostridium tetani , неправильная коллонизация после терапии антибиотиками и как следствие выработка нейротоксинов, а также значительное увеличение липополисахаридов (ЛПС) в крови у детей с аутизмом (Mangiola F. et al., 2016).
В случае депрессии — к этому времени причинами рассматривались нервно-психические расстройства, иммунологические процессы, генетическая предрасположенность или факторы окружающей среды. В последнее время растет научный интерес к корреляции с кишечным микробиомом. В клинических исследованиях, в которых характеризовался микробиом кишечника у пациентов с симптомами депрессии и тревожности отмечалось скопление выше среднего Alistipes из штамма Bacteroidetes, Lactobacillus spp . и бифидобактерий.
Изменения микробиома вызывают повышение проницаемости слизистой оболочки кишечника и, таким образом, увеличение ЛПС в крови, что приводит к активизации воспалительных процессов и, наконец, считается причинным фактором при нервно-психических заболеваниях.
При Faecalibacterium prausnitzii с другой стороны, оказывалась негативная корреляция между относительной частотой микробов и тяжестью проявления депрессии. Oscillibacter spp., Микроб, относящийся к классу клостридий, образует валерьяновую кислоту как основной метаболит. Валериановая кислота по структуре похожа на γ-аминомасляной кислоты (GABA) и, как было показано, может связываться с рецептором GABAa. Считается, что бактерии, участвующих в производстве или метаболизме валериановой кислоты, могут быть связаны с депрессией.
Метагеномный анализ кала для выявления дисбактериоза включает молекулярно-генетический анализ всего бактериального микробиома кишечника — в отличие от обычной диагностики кала, с помощью которой можно идентифицировать лишь ограниченное долю существующих кишечных микробов. Данные о микробиоме и их оценки регулярно обновляются и расширяются, чтобы новые результаты нашли отражение в следующих выводах.
________________________________
Рис. 10: Тестовый набор «Кишечный микробиом»
Стандарт микробиологической диагностики кала в 20 веке базировался прежде всего на культуральной системе, основанной на питательных средах и дальнейшей ферментативной дифференциации. Однако при культуральных методах вырастает очень мало анаэробных бактерий (примерно 99% кишечных бактерий). В последние годы методы исследования быстро развивались. В современных лабораториях идентификация бактерий в питательных средах в основном производится с помощью масс-спектрометрии (MALDI-TOF = Матрично-активированная лазерная десорбция / ионизация) или непосредственно из полученного материала с помощью молекулярно-биологических методов (PCR = полимеразная цепная реакция). Однако невыгодным является то, что ПЦР — это очень селективная и затратная технология.
Молекулярно-генетическое обследование « Микробиом кишечника»включает:
- Анализ биоразнообразия микробиома кишечника , включая грибы и дрожжи.
- рН-значение
- Приобретенный энтеротип
- Определение соотношения Firmicutes / Bacteroidetes ( индекс ожирения)
- Оценка Akkermansia muciniphila и провоспалительных бактерий (защитная флора слизистой оболочки)
- Заключение о дисбактериозе
- Анализ факторов риска заболеваний, связанных с микробиомом
- Интерпретация результатов
- Рекомендации по терапии, включая советы по питанию
Рис. 11: Образец результатов «Микробиом», 1 страница
Literatur Abell G. C. et al. Phylotypes related to Ruminococcus bromii are abundant in the large bowel of humans and increase in response to a diet high in resistant starch. FEMS Microbiol Ecol. 2008, 66(3):505-15. 2.Anhê F. F. et al. A polyphenol-rich cranberry extract preotects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut. 2014, 0:1-12. 3.Anonye B. O. Commentary: Dietary Polyphenols Promote Growth of the Gut Bacterium Akkermansia muciniphila and Attenuate High-Fat Diet-Induced Metabolic Syndrome. Front Immunol. 2017, 8:850. 4.Bakker-Zierikzee A.M. et al. Effects of infant formula containing a mixture of galacto- and fructo-oligosaccharides or viable Bifidobacterium animalis on the intestinal microflora during the first 4 months of life. Br J Nutr. 2005, 94(5):783-90. 5.Cardona F. et al. Benefits of polyphenols on gut microbiota an implications in human health. J Nutr Biochem. 2013, 24(8):1415- 1422. 6.Cuervo A. et al. Pilot study of diet and microbiota: interactive associations of fibers and polyphenols with human intestinal bacteria. J Agric Food Chem. 2014, 62(23):5330-6. 7.Costabile A. et al. A double-blind, placebo-controlled, cross-over study to establish the bifidogenic effect of a very-long-chain inulin extracted from globe artichoke (Cynara scolymus) in healthy human subjects. Br J Nutr. 2010, 104(7):1007-17. 8.Davis L. M. G. et al. Barcoded Pyrosequencing Reveals That Consumption of Galactooligosaccharides Results in a Highly Specific Bifidogenic Response in Humans. PLoS ONE, 2011, 6(9):e25200. 9.Drabińska N. et al. The Effect of Oligofructose-Enriched Inulin on Faecal Bacterial Counts and Microbiota-Associated Characteristics in Celiac Disease Children Following a Gluten-Free Diet: Results of a Randomized, Placebo-Controlled Trial. Nutrients. 2018, 10(2):201. 10.Everard A. et al. Microbiome of prebiotic-treated mice reveals novel targets involved in host response during obesity. The ISME Journal. 2014, 8:2116-2130. 11.Henning S. M. et al. Decaffeinated green and black tea polyphenols decrease weight gain and alter microbiome populations and function in diet-induced obese mice. Eur J Nutr. 2018, 57(8):2759-2769. 12.Hustoft T. N. et al. Effects of varying dietary content of fermentable short-chain carbohydrates on symptoms, fecal microenvironment, and cytokine profiles in patients with irritable bowel syndrome. Neurogastroenterol Moil. 2017, 29(4). 13.Kleessen B. et al. Jerusalem artichoke and chicory inulin in bakery products affect faecal microbiota of healthy volunteers. Br J Nutr. 2007, 98(39):540-9. 14.Lee H. C. et al. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res Microbiol. 2006, 157(9):876-84. 15.Moro G. et al. Dosage-related bifidogenic effects of galacto- and fructooligosaccharides in formula-fed term infants. J Pediatr Gastroenterol Nutr. 2002, 34(3):291-5. 16.Ramirez-Farias C. et al. Effect of inulin on the human gut microbiota: stimulation of Bifidobacterium adolescentis and Faecalibacterium prausnitzii. Br J Nutr. 2009, 101(4):541-50. 17.Roopchand D. E. et al. Dietary polyphenols promote growth of th gut bacterium Akkermansia muciniphila and attenuate high-fat diet-induced metacbolic syndrome. Diabetes, 2015, 64:2847-2858. 18.Sánchez-Patán F. et al. In vitro fermentation of a red wine extract by human gut microbiota: changes in microbial groups and formation of phenolic metabolites. J Agric Food Chem. 2012, 60(9):2136-47. 19.Singh R. K. et al. Influence of diet on the gut microbiome and implications for human health. J Transl Med. 2017, 15:73. 20.Tzounis X. et al. Flavanol monomer-induced changes to the human faecal microflora. Br J Nutr. 2008, 99(4):782-92. 21Venkataraman A. et al. Variable responses of human microbiomes to dietary supplementation with resistant starch. Microbiome. 2016, 4(1):33. Viebahn I. et al. Effekte einer prebiotischen Intervention auf Darmbeschwerden, Wohlbefinden und Mikrobiom-Diversität. 2018. Vivatvakin, B. et al. Effect of a whey-predominant starter formula containing LCPUFAs and oligosaccharides (FOS/GOS) on gastrointestinal comfort in infants. Asia Pac J Clin Nutr. 2010, 19(4):473-80. Zhu, L. et al. Inulin with different degrees of polymerization modulates composition of intestinal microbiota in mice. FEMS Microbiology Letters. 2017, 364
причины, симптомы и пути лечения
Слизистая оболочка верхних дыхательных путей представляет собой первую линию защиты организма против разнообразных патогенных факторов окружающей среды, таких как бактериальные, грибковые, вирусные, промышленные химические раздражители и загрязнения и обеспечивает противовирусный иммунитет. Повреждение слизистой или сухость облегчают проникновение вируса в организм. После того, как враг попал в организм и начал повреждать клетки, огромное значение играет выработка интерферонов, которые обеспечивают их невосприимчивость к действию вируса.
Противовирусный иммунитет – это очень сложный процесс, в котором участвует множество ресурсов иммунной системы. К сожалению, иммунные реакции не всегда срабатывают так, как об этом пишут в учебниках по биологии. Часто какой-либо процесс может быть нарушен, что приводит к осложнениям и проблемам. Когда снижен иммунный ответ, нужны средства, поднимающие иммунитет. Они могут быть природными, либо купленными в аптеке, главное – это эффективность и безопасность.
Слизистая оболочка верхних дыхательных путей большей частью покрыта тонким специализированным эпителием, создающим восприимчивый (чувствительный) барьер, который постоянно бомбардируется экзогенным живым или мертвым антигенным материалом.
Вдыхаемый воздух содержит значительное количество потенциально вредных веществ (газы, взвешенные корпускулярные частицы, бактерии, вирусы, микоплазмы, грибы, в том числе дрожжи, риккетсии, пыльцу, частицы слущенного эпителия человека и животных и др.). Согласно концепции местного иммунитета, слизистые оболочки и кожа как покровы, обращенные во внешнюю среду, защищают внутреннюю среду организма и сохраняют постоянство внутренней среды путем тесного взаимодействия эволюционно выработанного комплекса неспецифических и специфических механизмов защиты.
Местный иммунитет ЛОР-органов — это барьерная функция слизистых оболочек, включающая лимфоэпителиальные органы глотки, расположенные на пересечении воздухоносных и пищепроводных путей, первыми реагирующие на очередное антигенное раздражение (инфекцию) включением механизмов иммунной защиты, и неспецифические защитные факторы слизистой оболочки (мукоцилиарный транспорт, продукция лизоцима, лактоферрина, интерферона и др.)
А теперь непосредственно о микрофлоре верхних дыхательных путей и полости рта. Важно понимать, что ЛОР-органы не являются исключением в отношении микробной стерильности, т. е. в норме мы можем выявлять определенные микроорганизмы в носу, глотке и наружном ухе. Эти микробы в условиях сохранения иммунной резистентности не только не приносят нам никакого вреда, но и наоборот, оказывают положительное влияние. Они образуют так называемую биопленку (толщиной от 0,1 до 0,5 мм) – полисахаридный каркас которой, состоящий из микробных полисахаридов и муцина, предотвращает заселение организма патогенными микроорганизмами. Благодаря выделению микроорганизмами биопленки различных кислот, спирта, лизоцима (антибактериальное вещество) и стимуляции образования иммуноглобулина А тормозится развитие патогенных микробов. Кроме того, нормальные микроорганизмы препятствуют выделению токсинов патогенными бактериями.
Иллюстрацией работы «полезных» микроорганизмов является, например, следующий факт: с каждым вдохом мы в среднем вдыхаем около 1400-1700 микробов, а благодаря, в частности, биологическим факторам неспецифической защиты в глотке их количество сокращается до 300, в гортани и нижележащих отделах эти микроорганизмы не обнаруживаются вообще.
Это лишний раз подтверждает важность сбалансированной микрофлоры для полноценной жизнедеятельности всего организма.
Нормальная микрофлора глотки разнообразна, поскольку здесь смешивается микрофлора полости рта и воздухоносных путей. Представителями резидентной микрофлоры считаются: нейссерии, дифтероиды, и гемолитические стрептококки, энтерококки, микоплазмы, коагулазоотрицательные стафилококки, моракселлы, бактероиды, боррелии, трепонемы, актиномицеты. В верхних дыхательных путях преобладают стрептококки и нейссерии, помимо этого встречаются стафилококки, дифтероиды, гемофильные бактерии, пневмококки, микоплазмы, бактероиды. Слизистая оболочка гортани, трахеи, бронхов в норме стерильна. Следует сказать, что вышеуказанный видовой спектр микроорганизмов не является неизменным. Его состав зависит от многих факторов: сопутствующей патологии, возраста, условий внешней среды, условий труда, питания, перенесенных заболеваний, травм и стрессовых ситуаций и т.д.
Такие состояния, как острые и хронические заболевания всего организма, а особенно уха, горла, носа и полости рта, аллергия, неблагоприятные физические факторы (переохлаждение, инсоляция и т. д.), химические агенты, физическое, интеллектуальное и психоэмоциональное перенапряжение, прием антибактериальных, гормональных и цитостатических препаратов, недостаточное питание, гиповитаминоз, профессиональные вредности, курение, алкоголь провоцируют нарушение нормального микробиологического равновесия в ЛОР-органах и являются причинами нарушения качественного и количественного состава микрофлоры ротоглотки. Существует бактериологическое понятие, которое характеризует изменение соотношения представителей нормальной микрофлоры, снижение числа или исчезновение некоторых видов микроорганизмов за счет увеличения количества других и появления микробов – это дисбактериоз.
Терапия дисбактериоза должна быть строго индивидуальной и направленной в первую очередь на ликвидацию источника проблемы. Например, пока не будет адекватно пролечен аденоидит (консервативно или оперативно), от дисбактериоза вряд ли возможно избавиться. Вопрос о необходимости приема антибиотика определяется исключительно течением основного заболевания: если дисбактериоз глотки вызван наличием у ребенка хронического панкреатита, то антибактериальные средства тут неуместны; если же дисбактериоз связан с запущенной формой хронического синусита или аденоидита – без этой группы лекарственных препаратов, скорее всего, не обойтись. Воспалительные очаги в носо- и ротоглотке должны быть обязательно устранены – кариозные зубы пролечены, аденоидные вегетации, небные миндалины санированы и т.д. Помните, что они могут не только вызывать местные воспалительно-инфекционные процессы и дисбактериоз ротоглотки, но и являться источником аутоинфекции и аутоинтоксикации всего организма. Убедительные данные свидетельствуют, что возникновение ряда заболеваний респираторной, желудочно-кишечной, сердечно-сосудистой систем, опорно-двигательного аппарата обусловлено очаговой инфекцией в ротоглотке. Поэтому санация полости рта и ЛОР-органов – это обязательное условие и первый шаг к выздоровлению при многих патологических процессах.
Не стоит также забывать о мерах профилактики развития инфекционно-воспалительных процессов ротоглотки и ЛОР-органов. Так, стандартными мерами профилактики являются: ежедневные гигиенические мероприятия (туалет носа), регулярное посещение врача, рациональное питание, закаливание, создание оптимального микроклимата (влажность, температура), лечение сопутствующей патологии.
Кроме вышеперечисленных мер профилактики, помочь решить проблему инфекционно-воспалительных заболеваний ЛОР-органов и помочь восстановить нормальный микробиоценоз слизистой оболочки может прием средств растительного происхождения на основе прополиса.
Прополис – это смесь природных смол, вырабатываемых пчёлами из соединений, собираемых с разных частей растений, бутонов и выделений. Современные противомикробные применения прополиса включают рецептуры для лечения синдрома простуды (инфекции верхних дыхательных путей, обычная простуда и гриппоподобные инфекции), для заживления ран, лечения ожогов, акне, простого и генитального герпеса, и нейродермита.
В настоящее время, прополис является природным лечебным средством, он применяется и в косметике и является популярным альтернативным лекарственным средством для самолечения различных заболеваний. Современное применение прополиса предполагает его использование для лечения синдрома простуды (инфекции верхних дыхательных путей, обычная простуда и гриппоподобные инфекции), а также в качестве дерматологических препаратов, полезных для заживления ран, лечения ожогов, акне, простого и генитального герпеса, и нейродермита. Прополис также применяют для полоскания рта для предупреждения кариеса и для лечения гингивита и стоматита. На рынке он представлен в комбинации с экстрактом тимьяна, меда и витамина С в форме спрея для полости рта и горла Санорин Лорис, который не содержит спирта и является водно-глицериновым раствором. Благодаря комбинации натуральных компонентов и их свойствам, спрей Санорин Лорис эффективен для гигиены полости рта и для профилактики раздражения, увлажнения и защиты слизистой горла, а также как дополнение к комплексному лечению и профилактике ОРЗ. Важно отметить, что это удобный спрей для использования в любой ситуации, достаточно встряхнуть флакон и направить распылитель на проблемный участок.
Bacteroides — обзор | ScienceDirect Topics
Bacteroides
Bacteroides — это род грамотрицательных, неспорообразующих, облигатно анаэробных палочковидных бактерий. Распознано более 30 видов Bacteroides . Строгое таксономическое определение Bacteroides ограничивает эту перепись менее чем дюжиной отдельных видов. В семействе Bacteroidaceae организмов Bacteroides отличаются гуанин-цитозиновым составом ДНК — от 40 до 48 мол.%.Основными продуктами их сахаролитического метаболизма являются ацетат, сукцинат и изовалерат. Насыщенные антеизо-метил и изометил-разветвленные кислоты используются для их идентификации на основе длинноцепочечных жирных кислот. Bacteroides организмов экспрессируют ферменты гексозомонофосфатного шунта – пентозофосфатного пути. Их мембраны, богатые сфинголипидами, также содержат менахиноны, особенно MK-10 и MK-11. Bacteroides пептидогликан содержит мезо-диаминопимелиновую кислоту. Некоторые из Bacteroides spp.экспрессируют многочисленные капсульные полисахариды. Эти гликоантигены представляют биологический интерес из-за их иммуномодулирующего потенциала, особенно в случае полисахарида A. fragilis A. визуализируется при естественном освещении и в виде флуоресцентного излучения после воздействия ультрафиолетового (УФ) света и многочисленных биохимических тестов. Анализ короткоцепочечных жирных кислот с помощью газожидкостной хроматографии также можно использовать для дискриминации Bacteroides на уровне видов.Группа B. fragilis имеет особое медицинское значение по нескольким причинам. Часто это преобладающий GNAR при полимикробных инфекциях, и члены этой группы могут экспрессировать β-лактамазу. B. fragilis Бактериемия группы также связана с высокой смертностью: 27%. 2 Следовательно, идентификация группы B. fragilis клинической лабораторией часто имеет решающее значение для соответствующего терапевтического вмешательства. На кровяном агаре Б.fragilis образует круглые, цельные, белые или серые колонии размером 2-3 мм, блестящие и гладкие. Группа B. fragilis может быть довольно плеоморфной при окраске по Граму, образуя прямые палочки разной длины, а также коккобациллы (рис. 249-1A). При выращивании в жидкой среде в клетках образуются биполярные вакуоли, которые имеют характерный вид «английской булавки». Полезной характеристикой группы B. fragilis является ее толерантность к желчи по сравнению с другими GNAR, что позволяет ей расти на желчно-эскулиновом агаре Bacteroides .Кроме того, B. fragilis обладает высокой устойчивостью к антибиотикам канамицину, ванкомицину и колистину. Использование простого диско-диффузионного анализа для этих антибиотиков часто является частью процесса идентификации GNAR (таблица 249-1).
Бактероиды — обзор | Темы ScienceDirect
Bacteroides spp.Bacteroides видов — это грамотрицательные, анаэробные, устойчивые к желче бактерии, обитающие в кишечнике и составляющие примерно 25% кишечной микробиоты.Эти комменсальные бактерии могут влиять на иммунную систему кишечника, взаимодействуя с хозяином или производя определенные молекулы, которые в конечном итоге изменяют иммунный ответ кишечника. Различные штаммы Bacteroides были изучены на предмет их пробиотических свойств. Bacteroides fragilis продуцирует полисахарид А (ПСА), известный своими иммуномодулирующими свойствами, и активирует Т-клетки. Он также активирует TLR-2 PSA-зависимым образом, улучшая регуляторные Т-клетки для иммунной толерантности и поддержания гомеостаза кишечника [47, 48].
Б . fragilis Также сообщалось, что продуцирует фрагилизин, который считается фактором риска развития рака толстой кишки [49]. Эта функция не является желательной чертой для ее включения в список NGP. Около Bacteroides spp. также известны своей способностью расщеплять полисахариды растительного происхождения в кишечнике и производить промежуточные продукты для получения энергии. Bacteroides xylanisolvens, , например B . fragilis, , как было показано, увеличивает продукцию антител (IgM) против антигена Томсена-Фриденрейха (TFα) в зависимости от дозы и времени [50].Повышенное количество антител, специфичных к TFα, будет вызывать иммунный ответ хозяина для иммунного надзора против онкогенных клеток, присутствующих в системе. B. xylanisolvens считается одним из NGP из-за его противоопухолевых свойств без каких-либо доказательств токсичности [51].
Bacteroides uniformis, B. acidifaciens, и B. dorei упоминаются одинаково . Пероральное введение B. uniformis мышам, получавшим пищу с высоким содержанием жира, улучшило липидный профиль, уровни лептина и продукцию TNF-α DC, что связано с фагоцитозом [52]. Сообщалось, что B. uniformis улучшает иммунологические дисфункции и метаболические нарушения, связанные с дисбактериозом кишечника у мышей с ожирением. Острое введение этого штамма не показало каких-либо побочных эффектов.
Сообщалось также о том, что B. acidifaciens обладает потенциалом против ожирения, как предполагало исследование, проведенное на мышах с нокаутом [53]. Механизм предотвращения ожирения, вызванного диетой, заключается в стимулировании расхода энергии за счет активации пути TGR5 (мембранный рецептор желчной кислоты) –PPARγ в жировой ткани и увеличения уровня GLP-1 в плазме, что, вероятно, вызвано снижением уровня дипептидилпептидазы IV [ 53].
Возможное положительное влияние Bacteroides на активность липополисахаридов в фекалиях и сердечно-сосудистые заболевания
Ретроспективный анализ: дизайн исследования и участники
В нашем ретроспективном анализе мы получили доступ к 93 наборам клинических данных из двух независимых исследований (UMIN000015703 и UMIN000022414) (рис. ) 1,30 . Поскольку один пациент был исключен из-за отсутствия определяемых уровней ЛПС в фекалиях, мы наконец проанализировали данные 92 пациентов. Все участники исследования были набраны в университетской больнице Кобе в период с октября 2014 года по апрель 2017 года.Образцы фекалий были собраны, когда участники были госпитализированы и соблюдали больничную диету более 1 дня. Пациенты с сердечной недостаточностью поступали по поводу острой декомпенсированной сердечной недостаточности de novo или острого обострения хронической сердечной недостаточности. HF определялся на основе модифицированных критериев Фрамингема 31 . Пациенты с ИБС, перенесшие операцию по аортокоронарному шунтированию (АКШ) или чрескожное коронарное вмешательство, продемонстрировали стеноз> 75% при диагностической коронарной ангиографии, с сохраненной фракцией выброса левого желудочка и отсутствием СН в анамнезе.Исключались пациенты с острым коронарным синдромом. Пациенты контрольной группы, которые были госпитализированы в ту же больницу, имели гипертонию, сахарный диабет или дислипидемию в соответствии с рекомендациями 32 , но не имели в анамнезе ИБС и сердечную недостаточность. Пациенты, прошедшие курс лечения антибиотиками в течение месяца до поступления и во время госпитализации, были исключены. Все участники предоставили письменное информированное согласие, и исследование было проведено в соответствии с принципами Хельсинкской декларации. Это исследование было одобрено Комитетом по медицинской этике Университета Кобе (No.B1) и был зарегистрирован в Реестре клинических исследований Медицинской информационной сети университетской больницы (UMIN) (UMIN000036752).
Рисунок 1Обзор экспериментального плана. Это исследование состояло из трех отдельных исследований: исследование A, исследование B и исследование C. Целью исследования A было выяснить, какие и как конкретные кишечные бактерии влияют на уровень ЛПС в фекалиях, используя данные о людях. Целью исследования B было изучить биоактивность LPS кишечных бактерий, определенную в исследовании A in vitro и in vivo.Целью исследования C было выяснить, что пробиотики могут изменять кишечную флору пациентов до симбиотических состояний, снижая их уровень ЛПС в фекалиях до клинических испытаний (см. Рис. 1). ВНИМАНИЕ !!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!! CAD ишемическая болезнь сердца; E. coli , Escherichia coli ; HF сердечная недостаточность; LAL лизат амебоцитов Limulus; ЛПС липополисахарид.
Уровни ЛПС в фекалиях
Супернатант фекалий получали согласно ранее описанному протоколу с некоторыми модификациями 18 .Вкратце, образцы фекалий суспендировали в стерильном PBS до концентрации 1 г на 10 мл и слегка встряхивали, чтобы избежать разрушения бактериальных клеток. После центрифугирования в течение 15 минут при 3000 об / мин супернатант собирали и стерилизовали фильтрованием через фильтр 0,45 мкм с последующей повторной фильтрацией через фильтр 0,22 мкм и инактивацией в течение 15 минут при 90 ° C. Уровни ЛПС в фекалиях определяли с помощью анализа LAL (# K50-643 J; Lonza Inc., Базель, Швейцария) в соответствии с инструкциями производителя.Измерения ЛПС проводили в апирогенных стеклянных пробирках, пробирках Эппендорфа и 96-луночных планшетах.
Экстракция ДНК, амплификация гена 16S рРНК и пиросеквенирование
Собранные образцы фекалий хранили при -80 ° C. Экстракция ДНК из образцов фекалий человека была проведена Nihon Gene Research Laboratories Inc. в соответствии с ранее установленной процедурой 33 .
Части генов 16S рРНК (область V3 – V4, соответствующая положениям с 342 по 806 в клетке E.coli ) были амплифицированы с помощью ПЦР с использованием нашего невырожденного универсального набора праймеров 342F и 806R. Подробное описание набора праймеров и условий ПЦР доступно в другом месте 34 . После добавления адаптеров для секвенирования ампликоны секвенировали Takara Bio Inc. с использованием платформы Illumina MiSeq (Illumina Inc., Сан-Диего, Калифорния) в соответствии с протоколом производителя. Для создания матрицы бактериального состава использовали USEARCH версии 10.0.240. Наш предыдущий протокол также использовался для выбора последовательностей ампликонов гена 16S рРНК высокого качества, созданных с использованием Trimmomatic 35 версии 0.33 с параметрами «LEADING: 17 TRAILING: 17 AVGQUAL: 25 MINLEN: 100». Остальные операции чтения были обработаны с помощью команды -fastq_mergepairs программы USEARCH с параметрами по умолчанию. Затем мы удалили последовательности, в которых отсутствует праймерная область, с помощью Tagcleaner версии 0.16 с параметрами «-tag5 CTACGGGGGGCAGCAG -mm5 3 -tag3 AGATACCCCGGTAGTCC -mm3 3 -nomatch 3». После удаления праймера последовательности с неизвестными нуклеотидами (N) были удалены с помощью собственного скрипта Python. Чтобы удалить чтение PhiX, мы использовали команду -filter_phix для USEARCH.После этого мы удалили короткие последовательности (<300 нуклеотидов) с помощью команды USEARCH — sort_by_length, с параметром «-minseqlength 300». Наконец, мы сгенерировали таблицы операционных таксономических единиц (OTU) с помощью алгоритма UPARSE (команды -fastx_unique и otu_cluster с параметром «-minsize 1»). Репрезентативные последовательности каждой OTU были аннотированы для бактериального рода с использованием классификатора Ribosomal Database Project (RDP) версии 2.12 со значением начальной загрузки ≥ 0,5. Кроме того, мы аннотировали каждую репрезентативную последовательность каждой OTU в справочной базе данных Silva Living Tree Project, версия 123 36 , используя BLASTN версии 2.2.25, с порогом идентификации ≥ 97% и охватом ≥ 80%.
Животные
Мыши C57BL / 6J дикого типа были приобретены у CLEA Japan (Токио, Япония), а Tlr4 — / — мышей на фоне C57BL / 6J были приобретены у Charles River Japan (Йокогама, Япония). . Мышей содержали в специальном помещении, свободном от патогенов, в Институте Университета Кобе. Их кормили стандартной диетой (CE-2, CLEA, Токио, Япония) и водой ad libitum при строгом 12-часовом световом цикле.ЛПС E. coli , очищенный фенольной экстракцией, был приобретен у Sigma-Aldrich (Сент-Луис, Миссури, # L2630), и клетки Bacteroides были приготовлены для очистки ЛПС, как описано в Разд. «Экстракция ЛПС из Bacteroides vulgatus и B. dorei ». 8-недельным мышам дикого типа и Tlr4 — / — вводили ЛПС внутрибрюшинно для индукции септического шока 37 . Выживаемость контролировали ежедневно после инъекции LPS. Это исследование было одобрено Комитетом по этике животных Университета Кобе (P1
Экстракция ЛПС из
Bacteroides vulgatus и B. doreiLPS очищали методом горячего фенола, как описано ранее, с незначительной модификацией 38 . Бактерии культивировали в анаэробной среде Gifu (Nissui financial, Tokyo, Japan) при 37 ° C в течение 24 ч в анаэробных условиях. Затем 100 мл культивированных бактерий дважды промывали 15 мл PBS, содержащего 0,15 мМ CaCl 2 и 0,5 мМ MgCl 2 .Промытые клетки ресуспендировали в 8 мл PBS и разрушали обработкой ультразвуком на льду в течение 10 мин (модель Insonator 201 M; KUBOTA, Tokyo). После этого к суспензии клеток добавляли 80 мкл протеиназы К (10 мг / мл; Wako Pure Chemical, Осака, Япония) и инкубировали в течение 1 ч при 65 ° C. Затем следовала обработка ДНКазой I (20 мкг / мл; FUJIFILM Wako Pure Chemical, Осака, Япония) и РНКазой A (40 мкг / мл; Macherey-nagel, Дюрен, Германия) в присутствии 0,81 мМ MgSO 4 и 4 мкл / мл хлороформа при 37 ° C в течение ночи при встряхивании.К суспензии клеток добавляли равный объем фенола и встряхивали в течение 15 мин при 65 ° C. Полученную смесь центрифугировали (8500 × г, , 4 ° C, 15 мин), и водный слой собирали. К водному слою добавляли ацетат натрия до конечной концентрации 0,5 М. Затем добавляли этанол, в 10 раз превышающий объем водного слоя, и помещали при -20 ° C на ночь. Полученную смесь центрифугировали (2000 × г, , 4 ° C, 10 мин), и осадок ресуспендировали в воде. Остаточный фенол удаляли диализом против воды при 4 ° C с использованием Spectra / Por 6 (MWCO 1000; Spectrum Laboratories Inc., Ранчо Доминкес, Калифорния, США). После очистки материалы разделяли с помощью SDS-PAGE с использованием 4% стекирующего геля и 15% разделяющего геля и окрашивали серебряным пятном (Silver Stain Kit II, Wako Pure Chemical) и бриллиантовым синим кумасси для подтверждения чистоты. Сигналы детектировали с помощью ApeosPort-V C4475 (Fujixerox, Токио, Япония) с маркером молекулярного размера (EzStandard AE-1440, ATTO, Токио, Япония). Программное обеспечение для обработки изображений не использовалось. Очищенный ЛПС лиофилизировали и хранили при -20 ° C.
Анализы стимуляции цитокинов in vitro
RAW 264.7 макрофагов культивировали в среде RPMI-1640 с добавлением 10% фетальной бычьей сыворотки. Клетки (1 × 10 4 ) помещали в 96-луночные планшеты с плоским дном (Corning Costar, Corning, NY) и инкубировали в присутствии E.coli LPS или Bacteroides LPS в течение 12, 24 и более лет. 48 и 72 ч при 37 ° C с 5% CO 2 . Уровни цитокинов анализировали с использованием набора Cytometric Bead Array Kit (BD Biosciences, Сан-Диего, Калифорния, США) в соответствии с инструкциями производителя.
Культура фекалий в KUHIMM
Образцы фекалий были собраны у семи пациентов с ИБС во время их пребывания в больнице Университета Кобе в период с августа 2018 г. по май 2019 г. Каждый образец фекалий был собран с помощью тампона для анаэробной культуры (№ 212550; BD BBL, Нью-Джерси, США) и ферментировали в течение 24 часов 27 . Пациенты, прошедшие курс лечения антибиотиками в течение месяца до поступления и во время госпитализации, были исключены. Размер выборки был рассчитан с помощью программы R (мощность = 0.9, уровень значимости = 0,05, средняя разница = 8, стандартное отклонение = 5; N = 7 на группу). Все участники исследования предоставили письменное информированное согласие, и исследование проводилось в соответствии с принципами Хельсинкской декларации. Это исследование было одобрено Комитетом по медицинской этике Университета Кобе (№ 160191) и зарегистрировано в Реестре клинических испытаний UMIN (UMIN000024555).
Модель кишечной микробиоты человека Университета Кобе (KUHIMM) — это малогабаритный многоканальный ферментер (Bio Jr.8; ABLE, Токио, Япония), состоящий из восьми параллельных и независимых сосудов для анаэробного культивирования (рис. 1) 27,28,29 . Каждый сосуд содержал 100 мл анаэробной среды Gifu (Nissui Pharmaceutical Co, Токио, Япония) при pH 6,5. Культуры хранили при 37 ° C при постоянном перемешивании со скоростью 300 об / мин. Отфильтрованный газ N 2 : CO 2 (80:20) подавали непрерывно с постоянной скоростью потока 15 мл / мин для поддержания анаэробиоза. Образцы фекалий суспендировали в 2 мл 0,1 М фосфатного буфера (2: 1 NaH 2 PO 4 и 0.1 M Na 2 HPO 4 ) с добавлением 1% L-аскорбиновой кислоты. Сто микролитров фекальной суспензии засевали в один сосуд (культуральная группа). Сразу после инокуляции фекалий в сосуд добавляли по 1 × 10 8 клеток по B. vulgatus и B. dorei (группа Bacteroides). Фекальные культуры ферментировали в течение 48 часов.
Микробный анализ кишечных культур фекалий
Микробные профили кишечных микробов в образцах фекальных культур определяли секвенированием гена 16S рРНК, как подробно описано в другом месте 28,29 .Вкратце, область V3-V4 гена 16S рРНК была амплифицирована, и индексные праймеры были добавлены к ген-специфическим последовательностям. После ПЦР продукты очищали и элюировали 25 мкл 10 мМ Трис-буфера. Гены 16S рРНК и внутренний контроль (PhiX control v3; Illumina) подвергали 600 циклам секвенирования парных концов с использованием секвенатора MiSeq и набора реагентов v3 (Illumina, Inc.). Последовательности PhiX были удалены, и считывания парных концов с оценкой Q ≥ 20 были объединены с использованием программного обеспечения MacQIIME, версия 1.9.1 (Лаборатория Вернера, Кортленд, Нью-Йорк, США). Алгоритм UCLUST использовался для кластеризации отфильтрованных последовательностей в OTU на основе порога сходства ≥ 97%. Химерные последовательности были идентифицированы и удалены из библиотеки с помощью ChimeraSlayer. Репрезентативные последовательности из каждой OTU были таксономически классифицированы с использованием таксономической базы данных GreenGenes и классификатора RDP.
Статистический анализ
Статистический анализ выполняли с использованием Prism версии 7.0 (GraphPad Software; Сан-Диего, Калифорния), R версии 3.1.0 и JMP версии 14 (Институт SAS, Кэри, Северная Каролина). Для определения нормальности данных использовался тест Шапиро – Уилка. Результаты были выражены как среднее ± стандартная ошибка среднего для нормально распределенных данных и как медиана ± межквартильный размах (25–75-й процентили) для ненормально распределенных данных. Достоверность различий между двумя группами оценивалась с помощью двустороннего критерия Стьюдента t для нормально распределенных данных и U-критерия Манна-Уитни для ненормально распределенных данных.Различия в непрерывных параметрах между тремя группами рассчитывались с использованием одностороннего дисперсионного анализа для параметрических данных и теста Краскела – Уоллиса для ненормально распределенных данных. Выборки совпадающих пар сравнивали с использованием критерия знакового ранга Уилкоксона. Для сравнения категориальных переменных использовался точный критерий Фишера или критерий хи-квадрат. Кривые выживаемости Каплана-Мейера были построены и проанализированы с использованием лог-рангового критерия. Для всех тестов значение P <0,05 считалось показателем статистической значимости.Чтобы определить силу и направление связи между двумя параметрами, был рассчитан коэффициент ранговой корреляции Спирмена. Анализ главных компонентов и анализ дендрограммы выполняли с использованием программного обеспечения JMP версии 14 (Институт SAS). Модель случайного леса была построена с использованием пакета random Forest программы R.
Высокая распространенность устойчивости к клиндамицину у изолятов группы Bacteroides fragilis | Журнал антимикробной химиотерапии
Аннотация
Восприимчивость к антианаэробным агентам из группы Bacteroides fragilis варьируется в зависимости от изучаемого географического региона.В последние годы произошло снижение чувствительности таких изолятов, особенно к антибиотикам, таким как клиндамицин и цефокситин. Была определена противомикробная чувствительность 100 изолятов группы B. fragilis , выделенных в 1998 г. из фекалий здоровых людей, к клиндамицину и пяти другим антианаэробным агентам. Меропенем, метронидазол и тровафлоксацин показали отличную активность против всех изолятов. Эффективность цефокситина была низкой, только 46% изолятов были чувствительными.Наблюдалась высокая распространенность устойчивости к клиндамицину (49% изолятов).
Введение
Члены группы Bacteroides fragilis являются наиболее важными анаэробными грамотрицательными патогенами человека, являясь наиболее распространенным анаэробным организмом, поражающим кровоток. Они также являются наиболее распространенными анаэробными организмами, выделяемыми при внутрибрюшных инфекциях. 1 Восприимчивость к антианаэробным агентам варьируется в зависимости от географического региона и от одной больницы к другой в одном и том же регионе. 2 В последние годы произошло снижение чувствительности таких изолятов, особенно к определенным антибиотикам, таким как клиндамицин и цефокситин, при этом процент чувствительности составляет от 85 до 95% для B. fragilis и от 70 до 85% для другие представители группы B. fragilis . 1 Таким образом, информация о распространенности устойчивости в группе B. fragilis в каждом географическом регионе важна для правильного использования антианаэробных средств в профилактике или эмпирическом лечении инфекций, в которые могут быть вовлечены эти организмы.Мы изучили антимикробную чувствительность 100 изолятов группы B. fragilis к клиндамицину и пяти другим антианаэробным агентам: метронидазолу, меропенему, цефокситину, коамоксиклаву и тровафлоксацину.
Материалы и методы
Мы изучили 100 изолятов группы B. fragilis , выделенных из образцов стула, взятых у здорового населения административного медицинского района Мадрида, Испания, в 1998 г .; включая испытуемых обоих полов от 3 до 80 лет.Изоляты идентифицировали стандартными методами. МИК
определяли методом разведения на агаре Уилкинса – Чалгрена (Difco Laboratories, Детройт, штат Мичиган, США) для следующих антибиотиков: меропенема (Zeneca, Понтеведра, Испания), клиндамицина (Upjohn, Crawley, UK), метронидазола (Sigma , Сент-Луис, Миссури, США), цефокситин (Sigma), тровафлоксацин (Pfizer, Groton, CT, США) и коамоксиклав (SmithKline Beecham, Толедо, Испания).
Конечные концентрации составляли 0,5–256 мг / л клиндамицина, 0,12–8 мг / л меропенема, 4–64 мг / л цефокситина, 0.25–8 мг / л метронидазола, 1 / 0,5–16 / 8 мг / л коамоксиклава и 0,06–8 мг / л тровафлоксацина. Микроорганизмы инкубировали на кровяном агаре в течение 48 ч при 35 ° C в анаэробной камере для обеспечения чистоты и жизнеспособности. Планшеты MIC также инкубировали при 35 ° C в течение 48 ч в анаэробной атмосфере. В ходе исследования использовались штаммы для контроля качества B. fragilis ATCC 25285 и Bacteroides thetaiotaomicron ATCC 29741. Все результаты интерпретировали в соответствии с критериями NCCLS. 3
Результаты и обсуждение
MIC для двух контрольных штаммов всегда находились в рекомендуемых пределах. 3 Результаты исследования чувствительности суммированы в таблице, в которой показаны диапазоны, МИК 50 и МИК 90 , а также процентное соотношение чувствительных, промежуточных и устойчивых штаммов для каждого из протестированных антибиотиков.
Групповые инфекции B. fragilis требуют раннего лечения, обычно до получения результатов о чувствительности, особенно с учетом того факта, что тесты на чувствительность, используемые для этих микроорганизмов, более сложны и занимают больше времени, чем методы, обычно используемые для аэробных бактерий.Кроме того, в последние годы для группы B. fragilis сообщалось о повышении устойчивости к противомикробным препаратам, а также о региональных и институциональных различиях в показателях устойчивости. 2 , 4 Следовательно, результаты периодических исследований чувствительности полезны и необходимы в качестве руководства при выборе средств для профилактики и эмпирического лечения.
Результаты, полученные в этом исследовании, подтверждают превосходную активность in vitro метронидазола при 100% чувствительности штаммов и с очень низкими значениями МПК (≤0.5 мг / л) во всех случаях. Эти данные совпадают с данными предыдущих исследований 4 , 5 , хотя были описаны исключительные штаммы с устойчивостью к метронидазолу. 6 Меропенем также продемонстрировал высокую эффективность против группы B. fragilis , при этом резистентный штамм обнаружен не был. Устойчивость к карбапенемам регистрировалась нечасто. 4 Среди наших штаммов внимания заслуживают один с МПК 4 мг / л и четыре с МПК 2 мг / л.Эти высокие МИК, одно или два разведения ниже контрольной точки, должны привлечь внимание к прогрессивному снижению общей чувствительности к карбапенемам, ранее описанному другими авторами. 4 Похожая ситуация была обнаружена с коамоксиклавом. Хотя процент устойчивости в нашем исследовании (3%) соответствует описанному в предыдущей работе 7 , мы должны обратить внимание на частоту штаммов с промежуточной восприимчивостью (МПК 8/4 мг / л), достигающую 13%, и увеличение в MIC 90 , что точно совпадает с точкой останова.У наших изолятов не было обнаружено штамма с устойчивостью к тровафлоксацину. Проведено несколько предыдущих исследований, оценивающих чувствительность группы B. fragilis к тровафлоксацину. Хотя некоторые описывают низкий процент устойчивости, варьирующийся в зависимости от pH, 8 , большинство согласны с нашими результатами. 9
МИК, полученные для цефокситина, следует интерпретировать с осторожностью. Распределение MIC отображает режим, совпадающий с MIC, который NCCLS считает промежуточным по восприимчивости.Кроме того, 18 чувствительных штаммов имели МПК 16 мг / л, только одно разведение ниже контрольной точки, а 16 устойчивых штаммов имели МПК 64 мг / л, только одно разведение выше контрольной точки. В целом, 70 из 100 изолятов имели МИК, сгруппированные в трех разведениях, точка излома ± одно разведение. Это означает, что процентное соотношение резистентности и восприимчивости (18 и 46%) может измениться на 18 и 82% или на 54 и 46%, в зависимости от того, считаются ли штаммы с промежуточной восприимчивостью чувствительными или резистентными.Следовательно, хотя клиническое значение этой группы МИК , полученных in vitro , неизвестно, задействованные цифры делают использование этого агента нецелесообразным при инфекциях, вызванных группой B. fragilis .
Наиболее резкое увеличение показателей устойчивости по сравнению с предыдущими сообщениями было обнаружено при применении клиндамицина. Распространенность устойчивости к этому противомикробному препарату составила 49%, при МПК 90 256 мг / л. В отличие от цефокситина, распределение полученных МИК было бимодальным, так что было обнаружено, что подавляющее большинство изолятов явно чувствительны (≤0.5 мг / л) или явно резистентный (≥128 мг / л). Это повышение устойчивости к клиндамицину было описано ранее в нескольких отчетах, но только в одном, который был проведен в Испании в 1991 г., 7 , 10 были обнаружены схожими показателями устойчивости (45%). Другие современные или более поздние исследования в Испании 7 сообщают, что процент сопротивления ниже 25%. Географические различия очевидны. Европейское многоцентровое исследование показало, что резистентность колеблется от 0% в Австрии и Швеции до 19% в Бельгии. 2 Высокий процент устойчивости к клиндамицину в нашем исследовании имеет очевидные клинические последствия. Эндогенное происхождение инфекций, вызванных этим микроорганизмом, предполагает, что картина устойчивости, полученная в этом исследовании здоровой популяции, не сильно отличается от той, которая может быть обнаружена в тех клинических ситуациях, в которых участвует группа B. fragilis . Использование клиндамицина во многих учреждениях как часть эмпирического лечения абдоминальных или генитальных инфекций или его использование в протоколах хирургической профилактики в тех же областях должно быть изменено на основе повышения резистентности, как в нашем исследовании.Принимая во внимание большую географическую и даже институциональную изменчивость, описанную в предыдущих исследованиях, наши результаты должны служить для привлечения внимания к необходимости периодического обновления терапевтических протоколов в соответствии с результатами восприимчивости.
По нашему опыту меропенем, метронидазол и тровафлоксацин демонстрируют высокую эффективность и, таким образом, будут лекарствами выбора при инфицировании организмов группы B. fragilis . Коамоксиклав может быть хорошей альтернативой, несмотря на общую тенденцию к снижению восприимчивости.Цефокситин и клиндамицин демонстрируют заметное повышение уровня резистентности, и их использование в эмпирическом лечении инфекций, вызванных этими анаэробами, нецелесообразно. Их следует использовать только после получения результатов тестов на чувствительность. Таким образом, с учетом постоянно меняющихся моделей устойчивости к противомикробным препаратам группы B. fragilis , обновленная информация о моделях устойчивости и, как следствие, адекватная противомикробная терапия будет способствовать предотвращению терапевтических неудач в лечении инфекций, вызванных этим патогеном. .
Таблица I.Характер восприимчивости группы 100 Bacteroides fragilis , выделенных в 1998 г.
| . | МИК (мг / л) . | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| . | диапазон . | MIC 50 . | MIC 90 . | S (%) . | I (%) . | R (%) . | |||||||
| S, чувствительный; I — промежуточная восприимчивость; R, стойкий. | |||||||||||||
| Меропенем | ≤0,12–4 | 0,25 | 1 | 100 | 0 | 0 | |||||||
| Клиндамицин 256401 | 2 | 49 | |||||||||||
| Метронидазол | ≤0.25–0,5 | ≤ 0,25 | 0,5 | 100 | 0 | 0 | |||||||
| Цефокситин | ≤4–> 64 | 32 | 64 | 46 | 36401 Коамоксиклав | ≤1–16 | ≤1 | 8 | 83 | 13 | 4 | ||
| Тровафлоксацин | 0,12–2 | 0,25 | 0,5 | ||||||||||
| . | МИК (мг / л) . | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| . | диапазон . | MIC 50 . | MIC 90 . | S (%) . | I (%) . | R (%) . | ||||||||
| S, чувствительный; I — промежуточная восприимчивость; R, стойкий. | ||||||||||||||
| Меропенем | ≤0.12–4 | 0,25 | 1 | 100 | 0 | 0 | ||||||||
| Клиндамицин | ≤0,5–> 256 | 4 | 256 | 49 | 2 | 2 | ≤0,25–0,5 | ≤ 0,25 | 0,5 | 100 | 0 | 0 | ||
| Цефокситин | ≤4–> 64 | 32 | 64 | 46 | ||||||||||
| Коамоксиклав | ≤1–16 | ≤1 | 8 | 83 | 13 | 4 | ||||||||
| Тровафлоксацин | 0.12–2 | 0,25 | 0,5 | 100 | 0 | 0 | ||||||||
Характер восприимчивости 100 Bacteroides fragilis в группе, выделенной в 1998 г.
66 903 903 .| . | МИК (мг / л) . | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| . | диапазон . | MIC 50 . | MIC 90 . | S (%) . | I (%) . | R (%) . | ||||||||
| S, чувствительный; I — промежуточная восприимчивость; R, стойкий. | ||||||||||||||
| Меропенем | ≤0.12–4 | 0,25 | 1 | 100 | 0 | 0 | ||||||||
| Клиндамицин | ≤0,5–> 256 | 4 | 256 | 49 | 2 | 2 | ≤0,25–0,5 | ≤ 0,25 | 0,5 | 100 | 0 | 0 | ||
| Цефокситин | ≤4–> 64 | 32 | 64 | 46 | ||||||||||
| Коамоксиклав | ≤1–16 | ≤1 | 8 | 83 | 13 | 4 | ||||||||
| Тровафлоксацин | 0.12–2 | 0,25 | 0,5 | 100 | 0 | 0 | ||||||||
Ссылки
1Лорбер, Б. (1995). Bacteroides, Prevotella и Fusobacterium (и другие важные с медицинской точки зрения анаэробные грамотрицательные палочки). В «Принципы и практика инфекционных заболеваний» Манделла, Дугласа и Беннета, , 4-е изд. (Манделл Г. Л., Беннетт Дж. Э. и Долин Р., ред.), Стр. 2195–204. Черчилль Ливингстон, Нью-Йорк
2Филлипс, И., King, A., Nord, C.E. & Hoffstedt, B. (
1992
). Чувствительность к антибиотикам группы Bacteroides fragilis в Европе. Европейская исследовательская группа.Европейский журнал клинической микробиологии и инфекционных заболеваний
11
,292
–304,3Национальный комитет клинических лабораторных стандартов. (1998). Стандарты эффективности для испытаний на чувствительность к противомикробным препаратам — Восьмое информационное приложение: M100-S8 . NCCLS, Уэйн, Пенсильвания.
4Снидман, Д. Р., Макдермотт, Л., Кухурал, Г. Дж., Хехт, Д. У., Яннини, П. Б., Харрелл, Л. Дж. и др. . (
1996
). Анализ тенденций в моделях устойчивости к противомикробным препаратам среди клинических изолятов видов группы Bacteroides fragilis с 1990 по 1994 г.Клинические инфекционные заболевания
23, Прил. 1
,S54
–65,5Турген, П., Турген, В., Гурдо, М., Дюбуа, Дж. И Ламот, Ф. (
1994
).Продольное исследование чувствительности видов группы Bacteroides fragilis к пяти противомикробным препаратам в трех медицинских центрах.Противомикробные препараты и химиотерапия
38
,2276
–9,6Ротими, В. О., Хуршид, М., Брейзер, Дж. С., Джамал, В. Ю. и Ходахаст Ф. Б. (
1999
). Виды Bacteroides с высокой устойчивостью к метронидазолу: возникающая клиническая проблема?Клиническая микробиология и инфекция
5
,166
–9.7Бакеро, Ф. и Рейг, М. (
1992
). Устойчивость анаэробных бактерий к антимикробным препаратам в Испании.Европейский журнал клинической микробиологии и инфекционных заболеваний
11
,1016
–20,8Фалагас, М. Э., МакДермотт, Л. и Снидман, Д. Р. (
1997
). Влияние pH на чувствительность in vitro к противомикробным препаратам группы Bacteroides fragilis .Противомикробные препараты и химиотерапия
41
,2047
–9.9Wexler, H. M., Molitoris, E., Molitoris, D. & Finegold, S. M. (
1996
). In vitro активности тровафлоксацина против 57 штаммов анаэробных бактерий.Противомикробные препараты и химиотерапия
40
,2232
–5,10Пелаес, М. Т., Серценадо, Э., Родригес-Крейксемс, М. и Боуза, Э. (
1991
). Устойчивость анаэробных бактерий к антимикробным препаратам.Обзоры Инфекционные болезни
13
,183
.© 2000 Британское общество антимикробной химиотерапии
границ | Влияние вегетарианской и веганской диеты на микробиоту кишечника
Введение
Недавние исследования микробиома человека стали предметом всеобщего интереса. На протяжении десятилетий многие исследования выявили влияние микробиоты кишечника человека на физиологию организма-хозяина с новыми и неожиданно широкими последствиями для здоровья и болезней.
Микробиота человека, определяемая как совокупность всех таксонов микробов, связанных с людьми (бактерии, вирусы, грибы, простейшие, археи), состоит из недавно оцененных 3 × 10 13 (триллионов) микробов, обитающих на каждом человеке (1 ). Термин микробиом часто неправильно используется как синоним термина микробиота. Однако микробиом относится к каталогу этих микробов и их генов. Микробиом кишечника человека представляет собой ~ 3,3 миллиона неизбыточных микробных генов, что превосходит человеческий геном (около 21000 генов) в соотношении ~ 150: 1 (2).Интересно, что разнообразие микробиомов двух разных людей огромно по сравнению с их геномными вариациями человека; люди примерно на 99,9% идентичны друг другу с точки зрения их генома (3), но микробиом их кишечника может отличаться на 80–90% (4).
Последние достижения в области лабораторных методов позволили выявить функции микробиоты кишечника человека, связанные с иммунитетом, а также с желудочно-кишечной, мозговой и сердечно-сосудистой системами. Исследования также показали сильное влияние микробиоты кишечника человека на клетки-хозяева и гены.Это обширное взаимодействие позволило предположить, что микробиом эффективно функционирует как отдельный «орган».
Несколько исследований показали, что существует три основных бактериальных энтеротипа (5) (1) род Prevotella (считается в основном противовоспалительным и в других отношениях защитным), (2) род Bacteroides (более провоспалительный и, возможно, родственный к повышенному риску метаболического синдрома и других патологических состояний) и (3) род Ruminococcus (биологическое значение которого менее очевидно) (6).
Дисбаланс микробиоты кишечника был связан с желудочно-кишечными заболеваниями, такими как рефлюкс, пептические язвы, синдром раздраженного кишечника, неалкогольный стеатогепатит и воспалительные заболевания кишечника. Кроме того, некоторые системные состояния, такие как ожирение, атеросклероз, диабет 2 типа, рак, болезнь Альцгеймера и Паркинсона, боковой амиотрофический склероз, расстройство аутистического спектра, атопия и т. Д., Также, по-видимому, связаны с неблагоприятными изменениями в составе микробиоты кишечника (7-17 ).Накопление доказательств указывает на то, что микробиота кишечника является медиатором влияния питания на метаболический статус хозяина. Текущие исследования сосредоточены на установлении причинно-следственных связей между людьми и разработке терапевтических вмешательств, таких как индивидуальное питание (18).
Состав рациона, по-видимому, оказывает долгосрочное и острое воздействие на экосистему кишечной микробиоты (19, 20). Различные долгосрочные диеты, такие как вегетарианская / веганская диета против всеядности, оказывают значительное влияние на состав микробиоты кишечника.Показано, что различное содержание кишечной микробиоты обеспечивает различные метаболиты питательных веществ, называемых постбиотиками. Например, SCFAs, фитоэстрогены или изотиоцианаты больше связаны с пищей на растительной основе, а ТМАО и вторичные желчные кислоты — с диетой на основе мяса. Эти и другие постбиотики по-разному участвуют в метаболизме организма хозяина. Этот обзор будет сосредоточен на некоторых общих, а также на конкретных аспектах этой динамичной области исследований.
Микробиота кишечника: общие аспекты
Помимо бактерий, кишечник является хозяином множества царств: архей, вирусов и эукариот, включая виды грибов.Микробиота кишечника представлена более чем 1000 видами микробов, принадлежащих только к двум типам: Bacteroidetes и Firmicutes (21). На основе образцов стула человека в целом чаще всего встречаются роды Bacteroides, Prevotella, Bifidobacterium, Eubacterium, Clostridium, Streptococcus и Enterobacteriaceae . Следует отметить, что образцы стула дают разумные оценки микробиоты кишечника, а не полное представление (22).Это связано с тем, что анаэробные виды часто прикрепляются к слизистой оболочке кишечника, что затрудняет идентификацию всех видов бактерий, присутствующих в толстом кишечнике. Кроме того, вероятно, что биологическое значение любого рода или вида не определяется их относительной долей во всей экосистеме. Скорее, его значение наблюдается через его метаболизм / постбиотики и влияние на кишечную нервную систему, местный иммунитет, мозг и гены.
Влияние диеты на состав кишечной микробиоты
Разница в составе микробиоты кишечника между людьми, придерживающимися веганской / вегетарианской и всеядной диеты, хорошо задокументирована.Исследования показывают, что вегетарианские / веганские диеты способствуют развитию другой микробиоты по сравнению с всеядными, с незначительной разницей между веганами и вегетарианцами (23). Изменения в составе микробиоты могут быть связаны с различиями в бактериях, непосредственно потребляемых с пищей, различиями в потребляемых субстратах, вариациями времени прохождения через желудочно-кишечный тракт, pH, секрецией хозяина, на которую влияют режимы питания, и регуляцией экспрессии генов самого хозяина и / или или его / ее микробиота (24).
Растительная диета, по-видимому, полезна для здоровья человека, способствуя развитию более разнообразной микробной системы кишечника или даже распространению различных видов (25, 26).
Разнообразие и богатство кишечной микробиоты
Разнообразие микробиоты, по-видимому, имеет важную связь с ИМТ, ожирением и эластичностью артерий; и большинство исследований показывают, что диета на основе растений способствует большему микробному разнообразию. Клименко и др.обнаружили положительную связь между альфа-разнообразием или местным микробным богатством и долгосрочным потреблением фруктов и овощей ( p <0,05) (27). Аналогичным образом Martinez et al. наблюдали, что добавление цельнозернового ячменя, коричневого риса или смеси первых двух к рациону добровольцев привело к увеличению микробного разнообразия ( n = 28) (28). Клименко и др. также обнаружили отрицательную связь между альфа-разнообразием и ИМТ ( p <0,05) (27).
Однако кратковременное диетическое вмешательство с рекомендацией увеличить потребление клетчатки привело к небольшому, но значительному уменьшению разнообразия ( p <0.001). Исследователи предполагают, что это сокращение разнообразия может быть результатом быстрого изменения диеты, которое привело к временному нарушению микробного состава. Эта гипотеза преходящего микробного «стресса» также объясняет небольшое, но значительное увеличение содержания Enterobacteriaceae в результате вмешательства ( p <0,05). Enterobacteriaceae обычно ниже на веганской диете по сравнению с всеядной ( P <0,05) (29). Вероятно, это связано с большим присутствием бактерий, продуцирующих бутират, на диете с высоким содержанием клетчатки, что может снизить pH толстой кишки, предотвращая рост патогенных бактерий, таких как Enterobacteriaceae (30).
Verdam et al. наблюдали снижение микробного разнообразия у участников исследования с ожирением и без ожирения ( n = 28). Участники с ожирением также показали снижение соотношения Bacteriodetes : Firmicutes и увеличение Proteobacteria , провоспалительного типа. Аналогичным образом наблюдалось увеличение С-реактивного белка ( p <0,001), что обратно коррелировало с соотношением Bacteriodetes : Firmicutes ( p <0.05). Эти наблюдения предполагают провоспалительный эффект микробиоты, связанной с ожирением (31). С другой стороны, участники Adventist Health Study-2 (60 903), соблюдающие веганскую диету, показали самые низкие значения ИМТ по сравнению с теми, кто придерживался вегетарианской или всеядной диеты (32). Эти результаты показывают, что веганская диета, связанная с более низкой массой тела, может принести пользу микробному разнообразию и защитить от воспалений.
Menni et al. наблюдали, что скорость каротидно-бедренной пульсовой волны, мера жесткости артерий, отрицательно связана с разнообразием микробиома ( p = 0.001) у женщин ( n = 617) (33). Эта корреляция оставалась значимой после поправки на инсулинорезистентность и висцеральный жир. Жесткость артерий часто вызывается гипергликемией или гиперинсулинемией (34) и значительно коррелирует с воспалительными уровнями адипокинов. Исследователи предполагают, что связь между жесткостью артерий и микробным разнообразием можно частично объяснить ролью кишечника в модулировании системного воспаления. Таким образом, увеличение микробного разнообразия может улучшить системное воспаление, тем самым снижая жесткость артерий.
Кроме того, веганы и вегетарианцы имеют значительно большее богатство (альфа-разнообразие) по сравнению с всеядными животными, в частности, количество определенных операционных таксономических единиц (OTU), связанных с Bacteroidetes (35). Кажется вероятным, что многие преимущества вегетарианской / веганской диеты для здоровья частично опосредованы микробиотой кишечника, а не только за счет более высокой относительной численности тех ОТЕ, которые в настоящее время считаются защитными ( Bacteroidetes, Prevotella, Roseburia и т. Д. .), но также от постбиотических и эпигенетических воздействий на различные факторы риска хронического воспаления и хронических дегенеративных заболеваний (36).
Влияние диеты на Bacteroidetes: Firmicutes Ratio
Несмотря на значительные межиндивидуальные различия, здоровый микробиом кишечника взрослого человека характеризуется преобладанием этих Bacteroidetes -связанных ОТЕ наряду с таковыми из филума Firmicutes (37, 38). Исследования показали, что на изменчивость концентраций этих типов сильно влияет диета, в частности, соотношение между ними при сравнении всеядных диет типа, распространенного в Северной Америке, и диеты.вегетарианская / веганская диета. В одном исследовании сравнивали бактериальный состав взрослых индийцев ( n = 11) и китайцев ( n = 5) (39). В то время как обе группы населения придерживались диеты, основанной на углеводах и овощах, китайская диета была более тяжелой по животным жирам и белкам, чем индийская диета из цельнозерновых и растительных вегетарианских продуктов. Процент Bacteroidetes в микробиомах индийских участников был почти в четыре раза больше, чем у китайцев, 16.39% против 4,27% соответственно ( p = 0,001). Предполагается, что более высокая численность Bacteroidetes у индейцев связана с более низким потреблением ими продуктов животного происхождения; указывает на то, что диета с низким содержанием продуктов животного происхождения связана с большим количеством бактерий Bacteroidetes .
В другом исследовании сравнивали фекальную микробиоту итальянских детей ( n = 15) с фекальной микробиотой детей, живущих в сельских районах западной Африки, особенно в Буркина-Фасо ( n = 14) (40).Итальянские дети обычно придерживались западной диеты с высоким содержанием животного белка, сахара, крахмала и жира и низким содержанием клетчатки. Африканские дети Буркина-Фасо придерживались диеты с низким содержанием жиров и животных белков и богатой крахмалом, клетчаткой и растительным белком. Обилие Firmicutes было в два раза больше у итальянских детей, чем у детей Буркина-Фасо (63,7 против 27,3% соответственно). Обилие Bacteroidetes у итальянских детей было меньше, чем в два раза, чем у детей Буркина-Фасо (22.4 против 57,7% соответственно). Снижение уровней Firmicutes , обычно происходящее в пользу Bacteroidetes и Bifidobacteria , как видно в ответ на увеличение резистентных крахмалов, может быть полезным для профилактики и лечения ожирения (41). Хотя эти корреляции между диетой и составом микробиоты наблюдаются среди разных популяций, важно учитывать другие факторы, которые могут играть роль, такие как этническая принадлежность, генотипы хозяина, факторы окружающей среды и т. Д.
Исследования показали, что баланс Bacteroidetes и Firmicutes является важным маркером ожирения и более высокого ИМТ. В частности, уменьшенное соотношение Bacteroidetes : Firmicutes имеет сильную отрицательную корреляцию с ИМТ (r s = 0,59, P <0,001) (31). Возможное объяснение этой корреляции можно найти в наблюдении, что 20% -ное увеличение количества Firmicutes и соответствующее уменьшение численности Bacteroidetes связано с увеличением сбора энергии на 150 ккал / день, что приводит к увеличению веса со временем.Следовательно, повышенное соотношение Bacteroidetes : Firmicutes , как видно на растительной диете с высоким содержанием клетчатки, может привести к потере веса за счет уменьшения количества энергии, извлекаемой из рациона. Необходимы дальнейшие исследования, чтобы определить, связано ли увеличение сбора энергии с соотношением Bacteroidetes : Firmicutes , способствующим ожирению, или представляющим опосредованный хозяином адаптивный ответ для ограничения поглощения энергии (42).
Исследования также показали противоположные тенденции для Firmicutes и Bacteroidetes .В одном исследовании сравнивали детей в США, придерживающихся западной диеты, с детьми из Бангладеш, потребляющими растительную диету, состоящую из риса, хлеба и чечевицы. Соотношение Bacteroidetes : Firmicutes было в три раза выше у американских детей, потребляющих западную диету (43). Это противоречит предыдущему предсказанию западной диеты, приводящей к уменьшению соотношения Bacteroidetes : Firmicutes . Исследователи отметили возрастные и географические различия как потенциальное объяснение этого отклонения от ожидаемого соотношения, а также межпредметную изменчивость.В другом исследовании участникам предлагалось увеличить потребление клетчатки и избегать диетических продуктов западного происхождения. В то время как предыдущие исследования предполагали увеличение Bacteroidetes : Firmicutes , соотношение уменьшилось (от 0,13 ± 0,2 до 0,03 ± 0,09, парный тест Вилкоксона p <0,0001, n = 430) (27). Другое исследование проанализировало микробный состав худых и полных субъектов ( n = 98) и обнаружило, что по сравнению с худыми добровольцы с избыточным весом и ожирением имели более высокое соотношение Bacteroidetes к Firmicutes ( P = 0.001 и P = 0,005 соответственно) (44). Аналогичным образом, сравнение бактериальных типов не показало значительных различий в соотношении Bacteroidetes к Firmicutes между тучными и худыми добровольцами ( n = 20) (45). Эти примеры отражают трудности в широкой привязке определенных типов к конкретным диетам. Основная проблема при анализе конкретной микробиоты — это необходимость учитывать состояние и динамику взаимодействия микробов, охватывающих весь микробиом.
Влияние диеты на энтеротипы
Как упоминалось выше, в микробиомах человека наблюдаются три основных энтеротипа: Prevotella, Bacteroides и Ruminococcus . Prevotella , представитель филы Bacteroidetes , по-видимому, значительно богаче при соблюдении веганской диеты. В ранее упомянутом исследовании De Filippo et al., Prevotella присутствовала исключительно у детей Буркина-Фасо, потребляющих диету с низким содержанием жира и животного белка и богатую крахмалом, клетчаткой и растительным белком по сравнению с детьми, живущими в Италии употребление западной диеты с высоким содержанием животного белка, сахара, крахмала и жиров и низким содержанием клетчатки (40).В другом исследовании сравнивали диеты 178 пожилых жителей, живущих либо в общине, либо в учреждениях длительного ухода (46). Было обнаружено, что общественная группа придерживается диеты с низким и средним содержанием жиров и высоким содержанием клетчатки; в то время как жители, находящиеся под долгосрочным уходом, придерживались диеты с умеренным или высоким содержанием жиров и с низким содержанием клетчатки. Исследование показало, что у тех, кто в сообществе, профиль питания больше соответствует растительной диете, чаще встречаются кишечные сообщества энтеротипа Prevotella .
Исследование, в котором сравнивали взрослых людей из Индии и Китая, показало аналогичные результаты (39).Как и ожидалось, индейцы, которые потребляли меньше продуктов животного происхождения и больше продуктов растительного происхождения, чем китайцы, имели значительно больший процент Prevotella (13,07 против 0,58%, соответственно). Когда численность Prevotella была проанализирована у тайских вегетарианцев по сравнению с невегетарианцами, было обнаружено, что у вегетарианцев значительно выше количество Prevotella ( p = 0,005) (47). Другие исследования показали, что веганская / вегетарианская диета с высоким содержанием растительной пищи связана с высоким содержанием Prevotella (48, 49).Это предполагает дополнительную поддержку большего присутствия Prevotella у тех, кто потребляет меньше продуктов животного происхождения и больше продуктов растительного происхождения. В то время как исследования на мышах предполагают, что Prevotella улучшает метаболизм глюкозы за счет улучшения хранения гликогена (50), в настоящее время отсутствие каких-либо дополнительных исследований делает Prevotella просто родом, описывающим общую экосистему кишечных бактерий человека, в первую очередь при растительной диете. .
Bacteroides , другой основной энтеротип и род филы Bacteroidetes , также, по-видимому, зависит от диеты, но иначе, чем Prevotella . Bacteroides положительно коррелирует с долгосрочными диетами, богатыми животным белком и насыщенными жирами (20, 27). Вероятно, это связано с их способностью переносить желчь, которая часто встречается в кишечной среде тех, кто потребляет продукты животного происхождения. В упомянутом ранее исследовании, сравнивающем детей в США, придерживающихся западной диеты, и детей в Бангладеш, потребляющих растительную диету, Bacteroides был основным родом детской микробиоты США. Высокие пропорции Bacteroides обнаружены в кишечнике людей, потребляющих западную диету, и противоположное обнаружено у тех, кто придерживается диеты с высоким содержанием клетчатки из фруктов и бобовых (27, 37, 43, 47, 48).
Ruminococcus — третий по значимости энтеротип, связанный с длительным употреблением фруктов и овощей. Виды этого рода специализируются на разложении сложных углеводов, таких как целлюлоза и резистентный крахмал, содержащихся в растительных продуктах (51). Эти микробы разрушают пищевые волокна, производя бутират, который действует как противовоспалительное соединение. Ruminococcus положительно связан с низким ИМТ и отрицательно с плохим липидным профилем (27).Точно так же обилие Rumminococcus было связано с более низкой эндотоксемией и более низкой жесткостью артерий, что является предиктором сердечно-сосудистого риска (33). Потребление грецких орехов было в значительной степени связано с обогащением Ruminococcus (38). Тем не менее, Ruminococcus также был связан с низким потреблением пищевых волокон студентами колледжей. Хотя эти микробы разлагают сложные углеводы, они также разрушают устойчивые крахмалы, содержащиеся в продуктах из очищенного зерна (52). Ruminococcus также может играть роль в превращении холина животного происхождения в триметиламин (ТМА) (53). Таким образом, на численность Ruminococcus влияют диеты как животного, так и растительного происхождения.
Влияние диеты на количество бактерий
Хотя Bacteroides может вызывать воспаление, и их концентрация связана с долгосрочным потреблением продуктов животного происхождения, исследование, анализировавшее 11 вегетарианцев, 20 веганов и 29 всеядных животных (49), обнаружило расхождение в этом обобщении.В дополнение к обнаружению Clostridium clostridioforme и Faecalibacterium prausnitzii , которые считаются защитными для здоровья, в более высокой относительной численности у вегетарианцев / веганов по сравнению с всеядными животными, в этих группах также наблюдалось более высокое содержание Bacteroides thetaiotaomicron . Это несоответствие в классификации численности бактерий при растительной диете и диете на основе животных не является редкостью. Clostridium кластера XIVa был обнаружен в более низком соотношении у вегетарианцев / веганов, в отличие от исследования, показывающего, что бактерии кластера XIVa Clostridium являются основным компонентом микробиоты кишечника у женщин-вегетарианцев (103).Таким образом, хотя можно сделать обобщения, некоторые подтипы родов будут выбросами. Это несоответствие некоторых типов бактерий в зависимости от диеты было признано в предыдущих обзорных статьях и объяснялось различными причинами, такими как разные методологии профилирования микробиома, разные генетические особенности хозяина, индекс массы тела, а также потребление красного вина и аспартама (54, 55 ). Все эти факторы могут влиять на нашу микробиоту. Следовательно, необходимы дальнейшие исследования, чтобы отделить их эффекты от эффектов, связанных с растительным или растительным действием.всеядная диета.
В совокупности пищевые привычки влияют на состав кишечной микробиоты. Хотя диетические изменения оказывают относительно быстрое воздействие (51) (в течение недели) на микробный состав и, следовательно, на его метаболиты, эти эффекты являются умеренными и обратимыми (24). Например, изменения микробиоты и иммунных параметров после трехмесячной вегетарианской диеты значительны, но не отражают степень изменений, которые происходят при длительной вегетарианской диете (56).
Как компоненты растительной пищи влияют на кишечную микробиоту.
Биодоступность питательных веществ
Недавно было обнаружено важное значение потребления пищевых питательных веществ с низкой биодоступностью. Более низкая биодоступность питательных веществ, обнаруженная в более крупных частицах пищи, неповрежденных стенках клеток растений и продуктах без термической обработки, означает, что большее количество питательных веществ попадает в нижнюю часть желудочно-кишечной системы, тем самым улучшая доставку питательных веществ к микробиоте кишечника (57). Это помогает поддерживать нормальное развитие и функционирование кишечной микробиоты (57). Современные западные диеты содержат больше продуктов, подвергнутых ультраобработке, и бесклеточных питательных веществ, или питательных веществ, не содержащих клетки.Эти компоненты легче всасываются в тонком кишечнике, лишая толстую кишку важных питательных веществ, что может изменить состав и метаболизм кишечной микробиоты (58). Было показано, что бесклеточная пища, например сахар, вызывает воспаление у младенцев, подростков, женщин детородного возраста и пожилых людей. Цельные растительные продукты обладают защитным действием, способствуя росту полезных бактерий, разлагающих клетчатку, в толстой кишке (58).
Углеводы
В отличие от перевариваемых углеводов, неперевариваемые углеводы, такие как резистентный крахмал и некоторые сахара, достигают толстой кишки, где они могут ферментироваться микробиотой кишечника для обеспечения энергии или выработки постбиотиков.Однако как усвояемые, так и неперевариваемые углеводы могут влиять на микробиоту кишечника. Было показано, что усвояемые углеводы из фруктов (например, глюкоза, сахароза и фруктоза) уменьшают содержание Bacteroides и Clostridia (54). Неперевариваемые углеводы наиболее последовательно увеличивают количество молочнокислых бактерий, Ruminococcus, E. rectale и Roseburia , а также уменьшают количество видов Clostridium и Enterococcus (54). Было показано, что как усвояемые, так и неперевариваемые углеводы увеличивают содержание Bifidobacteria , рода актинобактерий типа .
В исследовании сравнивали уровни Bifidobacteria в ответ на рандомизированный двойной слепой перекрестный след. Участники употребляли как стандартную формулу для энтерального введения, так и формулу с добавлением фруктоолигиосахаридов (ФОС) и клетчатки (59) в качестве единственного источника питания в течение 14 дней. ФОС и клетчатка являются формами углеводов, которые естественным образом содержатся в большом количестве в растительных продуктах — бананах, артишоках, луке и т. Д. Хотя объем прописанной смеси основывался на индивидуальных расходах энергии, эталонный показатель в 2000 калорий формулы ФОС / клетчатка обеспечивал 10 .2 г ФОС и 17,8 г клетчатки. Среднее суточное потребление сбраживаемых неперевариваемых углеводов оценивается в 10 г из инулина и ФОС (60). В это количество не входят блюда и продукты с добавлением инулина и ФОС, которые обычно добавляют дополнительно 3–10 г на порцию. Следовательно, 10,2 г ФОС в формуле вполне реально для употребления человеком. 17,8 г клетчатки в формуле также подходят для потребления человеком, так как среднее потребление мужчин и женщин в США составляет 18 г и 15 г соответственно.
Bifidobacterium — это род, продуцирующий бутират, который, как известно, играет защитную роль в кишечном барьере человека, обеспечивая защиту от патогенов и болезней. Когда участникам давали смеси с ФОС и клетчаткой, их количество Bifidobacteria увеличилось с 5,1 до 26,6% ( P = 0,003) через 14 дней. Когда формула давалась без FOS и клетчатки, Bifidobacteria увеличивалось только с 3,3 до 8,6% ( P = 0,073). Наблюдалась отрицательная корреляция между исходным уровнем Bifidobacteria и величиной бифидогенного эффекта, что указывает на то, что пациенты с более низким исходным количеством Bifidobacteria получают наибольшую пользу от фруктоолигосахаридов и потребления клетчатки.Напротив, высокое потребление холестерина из продуктов животного происхождения было тесно связано с более низким содержанием Bifidobacteria (прил. p = 0,008).
Хотя эти исследования показывают, что Bifidobacterium увеличивается в ответ на диету, богатую клетчаткой и высоким содержанием углеводов, другие исследования показали противоречивые результаты. Одним из важных факторов может быть потребление алкоголя, которое было тесно связано с более низким содержанием Bifidobacteria (прил. р = 0,006). Исследователи, сравнивавшие уровни Bifidobacteria у веганов, вегетарианцев и контрольной группы, обнаружили, что Bifidobacteria значительно ниже ( p = 0,002) в веганских образцах, чем в контрольной группе, соблюдающей стандартную всеядную диету. Разницы между веганами и вегетарианцами не наблюдалось (29). Другое исследование выявило более высокие уровни Bifidobacteria у мясоедов по сравнению с участниками, которые перешли на вегетарианскую диету в течение 4 недель после употребления смешанной западной диеты с высоким содержанием жира и мяса (58).Относительное снижение содержания Bifidobacterium у вегетарианцев и веганов можно объяснить относительным обилием других видов защитных бактерий, таких как Prevotella . Prevotella обладает противовоспалительным действием (40) и может снижать рост других бактерий, конкурируя за клетчатку в качестве энергетического субстрата (61).
Недавнее исследование in vitro выяснило специфический механизм действия углеводов, в частности выбранных пищевых волокон, на микробиоту кишечника.Исследование показало, что следующие волокна обладают различными пребиотическими эффектами: инулин, альфа-связанные галактоолигосахариды, бета-связанные галактоолигосахариды, ксилоолигосахариды из кукурузных початков и сахарного тростника с высоким содержанием клетчатки, а также бета-глюкан из овса (62). . Бета-глюкан индуцировал рост Prevotella и Roseburia с одновременным увеличением продукции пропионата SCFA. Инулин и все олигосахариды обладали сильным бифидогенным действием (62). Это исследование также показало, что все натуральные сахара, особенно неперевариваемые формы, такие как инулин и олигосахариды, повышают уровни SCFA (62).Эффекты пребиотиков различаются в зависимости от типа бактерий, которыми расщепляется каждое волокно. Это определяется бактериальной специфичностью, в которой определенные кластеры генов в бактериальном геноме определяют сахаролитические ферменты, которые могут продуцировать бактерии, и, следовательно, могут ли они метаболизировать пребиотический субстрат (63). Неперевариваемые углеводы не только действуют как пребиотики, способствуя росту полезных микроорганизмов, но также снижают продукцию провоспалительных цитокинов, концентрацию триглицеридов в сыворотке крови, общего холестерина и холестерина ЛПНП (54).Таким образом, неперевариваемые углеводы могут оказывать защитное действие против сердечно-сосудистых заболеваний и заболеваний центральной нервной системы.
Белки
В большинстве исследований отмечается, что потребление белка положительно коррелирует с микробным разнообразием (54). Однако животные и растительные белки по-разному влияют на микробиоту кишечника. Например, люди, потребляющие диету с высоким содержанием животного белка, из говядины, которая также богата жирами, демонстрировали меньшее количество бактерий, таких как Roseburia, Eubacterium rectale и Ruminococcus bromii , которые метаболизируют полисахариды пищевых растений (51).Популяции бактерий, которые увеличиваются в ответ на диету с высоким содержанием животного белка по сравнению с субъектами, потребляющими постную диету, обычно являются устойчивыми к желчи микроорганизмами, такими как Bacteroides и Clostridia (64). Кроме того, диета с высоким содержанием белка обычно ограничивает потребление углеводов, что может привести к уменьшению количества бактерий, продуцирующих бутират, и, следовательно, к провоспалительному состоянию и повышенному риску колоректального рака (65).
У людей, потребляющих гороховый белок, наблюдается увеличение полезных Bifidobacterium и Lactobacillus и снижение патогенных Bacteroides fragilis и Clostridium perfringens и, как следствие, повышение уровней SCFA в кишечнике (54).Точно так же белки растительного происхождения связаны с более низкой смертностью, чем белки животного происхождения (54).
Жиры
Текущие данные свидетельствуют о том, что как количество, так и качество потребляемых жиров значительно влияют на состав микробиоты кишечника (65).
Растительная диета обычно имеет низкое содержание жиров, что способствует развитию полезных Bifidobacteria в кишечной микробиоте человека. Жир, который поступает из веганской / вегетарианской диеты, состоит преимущественно из моно- и полиненасыщенных жиров, которые увеличивают соотношение Bacteroidetes : Firmicutes , а на уровне родов — молочнокислые бактерии, Bifidobacteria и Akkermansia. muciniphila (54).Орехи, особенно грецкие, улучшают микробиоту кишечника за счет увеличения Ruminococcaceae и Bifidobacteria и уменьшения Clostridium sp. кластера XIVa видов (38).
С другой стороны, насыщенные жиры, которые содержатся почти исключительно в животных источниках, увеличивают Bilophila и Faecalibacterium prausnitzii и уменьшают Bifidobacterium (54). Некоторые исследования сообщают, что это изменение активирует воспаление (индуцирует провоспалительные цитокины, такие как IL-1, IL-6 и TNF-α) и приводит к метаболическим нарушениям (66).Высокое потребление насыщенных и трансжиров, преимущественно содержащихся в западной диете, увеличивает риск сердечно-сосудистых заболеваний и снижает количество Bacteroidetes, Bacteroides, Prevotella, Lactobacillus ssp. и Bifidobacterium spp, и увеличивает Firmicutes (40, 67).
Было обнаружено, что полиненасыщенные жирные кислотыN-3 либо не приводят к изменению микробиоты, либо приводят к положительному увеличению содержания Bifidobacterium, Adlercreutzia, Lactobacillus, Streptococcus, Desulfovibrio и Verrucomicrobia ( Akkerhila 67).
полифенолы
Полифенолы или встречающиеся в природе растительные метаболиты (68) в растительной пище увеличивают численность Bifidobacterium и Lactobacillus , которые обеспечивают антипатогенный и противовоспалительный эффект и защиту сердечно-сосудистой системы (54). Общие продукты, богатые полифенолами, включают фрукты, семена, овощи, чай, какао-продукты и вино. Например, экстракты полифенолов из чая увеличивают содержание Bifidobacterium и Lactobacillus – Enterococcus spp., что затем приводит к увеличению продукции SCFA на микробиоте человека in vitro (69).
Влияние постбиотиков микробиома на здоровье человека
Исследования осей кишечник-мозг, кишечник-легкие и кишечник-печень подчеркивают важность микробиома для системного здоровья человека. Исследования отмечают изменения в центральной нервной химии, воспалительных состояниях легких и патогенезе неалкогольной жировой болезни печени с изменениями микробного состава (70–72). Механизм связи между этими органами проистекает из микробных продуктов и микробных метаболитов проглоченных питательных веществ.Эти продукты могут быть независимыми от диеты (например, липополисахариды, синтезированные рибосомами посттрансляционно модифицированные пептиды и т. Д.), Но здесь мы хотели бы описать несколько примеров хорошо известных диетозависимых метаболитов, таких как SCFA и другие. В зависимости от бактерий и расположения в кишечном тракте из разных пребиотиков и питательных веществ могут образовываться разные биоактивные молекулы (70, 73). Микробные метаболиты могут оказывать разнообразные положительные эффекты на здоровье, включая местное противовоспалительное и иммуномодулирующее действие, а также системные анти-ожирение, антигипертензивное, гипохолестеринемическое, антипролиферативное и антиоксидантное действие (74).Эти постбиотические эффекты являются результатом модуляции экспрессии генов, метаболизма и функционирования кишечника и зависят от состава микробиоты и субстратов, в значительной степени от диеты.
Короткоцепочечные жирные кислоты
SCFAs действуют как субстрат для поддержания эпителия толстой кишки и коррелируют с потреблением растительной пищи (56). Поддержание кишечного барьера предотвращает эндотоксемию и последующие воспалительные эффекты (75, 76). SCFAs ацетат, пропионат и бутират, в основном, являются микробными метаболитами ферментированной клетчатки и других углеводов, хотя небольшая часть может происходить из белков.Уровни этих метаболитов (и соответствующих эфиров) в кале положительно коррелируют с потреблением фруктов, овощей и бобовых. Таким образом, их уровень значительно повышается у людей, переходящих на растительную диету. Интересно, что увеличение SCFAs наблюдается, когда всеядные животные потребляют средиземноморскую диету, богатую фруктами, бобовыми и овощами (77).
В то время как определенные кишечные микробы предрасположены к продукции SCFA, известно, что разные бактерии продуцируют разные SCFA.Например, кишечные бактерии, такие как Akkermansia muciniphila, Bifidobacterium spp., Prevotella spp. И Bacteroides spp. производят ацетат; Bacteroides spp. производить пропионат; и Coprococcus продуцирует бутират (78). Больше всего бутират-продуцирующих бактерий находится в Clostridium Cluster XIVa, IV и XVI. Эти виды положительно коррелируют с потреблением растительной пищи и производят SCFAs, которые приносят несколько преимуществ для здоровья.
Защитная роль ацетата, пропионата и бутирата против различных типов заболеваний, таких как диабет 2 типа, воспалительные заболевания кишечника и иммунные заболевания, хорошо документирована. Например, было показано, что SCFAs способствуют иммунитету против патогенов (78) и являются важными компонентами для функции и созревания микроглии и контроля целостности гематоэнцефалического барьера (79). Другой эффект SCFAs — усиление термогенеза, предотвращение / лечение ожирения (80, 81). SCFAs служат энергетическими субстратами для колоноцитов, а также для организма в целом.Например, пропионат служит глюконеогенным субстратом в печени и кишечнике (78).
Микробные взаимодействия с диетическими полисахаридами и образующимися SCFAs являются важными энергетическими и сигнальными молекулами. Все более широко признается, что бактерии, продуцирующие бутират, и бутират, как таковой , могут быть полезны для здоровья человека (78). Было показано, что бутират играет ключевую роль в физиологии кишечника как основной источник углерода для колоноцитов. Он помогает регулировать критические функции кишечника, такие как перистальтика кишечника, выработка слизи, висцеральная чувствительность, эпителиальный барьер, иммунный гомеостаз и градиент кислорода слизистой оболочки (82, 83).Таким образом, пищевые волокна и углеводы могут влиять на деградацию SCFA, изменяя при этом численность связанных микробов. В совокупности диета, богатая клетчаткой, может принести пользу кишечнику, а также улучшить общее состояние здоровья.
Фитоэстрогены
Фитоэстрогены — это полифенолы растительного происхождения, которые взаимодействуют с рецепторами эстрогенов либо агонистами, либо антагонистами. Подавляющее большинство полифенолов попадает в кишечник, учитывая их биодоступность 1% (57). Защитные эффекты растительных полифенолов, особенно их противораковые, противовоспалительные и антиоксидантные эффекты, а также их связь со снижением риска сердечно-сосудистых заболеваний, ожирения, диабета, остеопороза и образования амилоида, наблюдались у людей (84–86) .Все больше данных показывает, что эти эффекты достигаются после биоактивации полифенолов микробиотой кишечника (87, 88). Несмотря на то, что полифенолы растений обладают защитным действием на здоровье человека, особенно в биоактивной форме, все же существует возможность побочных эффектов из-за сложности их действия и потенциальной индивидуальной изменчивости (89).
Хотя пока известны не все типы микробов, участвующих в метаболизме полифенолов, недавно было показано, что Bifidobacterium, Lactobacillus sp., Coriobacteriaceae, Clostridium sp., Bacteroides и Saccharomyces дрожжи участвуют в процессе превращения полифенолов в эквол, уролитины и энтеролигнаны (74, 88). Качественные и количественные пропорции продуцируемых уролитинов и эквола положительно коррелируют с действием фитоэстрогенов (88). Другие виды бактерий, такие как Coriobacteriaceae и Eubacterium , ответственны за различные превращения полифенолов (88).
Взаимодействие полифенолов и кишечной микробиоты является двунаправленным (90, 91). Кишечные бактерии производят микробные метаболиты из полифенолов, которые, в свою очередь, служат пребиотиками для кишечных бактерий. Эти метаболиты, особенно уролитины, способствуют росту Lactobacillus и Bifidobacterium (88).
Витамины
Микробиота кишечника имеет решающее значение для адекватного уровня витаминов в организме человека. Менахинон, фолиевая кислота, кобаламин и рибофлавин (например, витамины K, B9 и B2) производятся кишечными микробами (25).Различные бактерии обладают биосинтетическими свойствами для разных витаминов, например, Bifidobacterium для витаминов K, B 12 , биотин, фолат, тиамин, Bacillus subtilis и Escherichia coli для рибофлавина (92) и Lactobacillus для Lactobac. и другие витамины группы В (93). Анализ путей предсказанных метагеномов показал обогащение биосинтеза фолиевой кислоты у веганов по сравнению с всеядными животными (77).
Изотиоцианаты
Изотиоцианаты — это соединения глюкозинолатов, которые в основном содержатся в растениях, например в овощах семейства крестоцветных. Escherichia coli , некоторые Bacteroides , некоторые Enterococcus, Lactobacillus agilis , некоторые Peptostreptococcus spp. и Bifidobacterium spp. метаболизируют глюкозинолаты до изотиоцианатов, секретируя собственный фермент мирозиназу (94). Эти метаболиты обладают цитопротекторным и антиоксидантным действием за счет регуляции экспрессии генов, связанных с тонопластическими, атеросклеротическими и нейродегенеративными процессами (25).
Лиганды арил-углеводородного рецептора
Лиганды кишечных арилуглеводородных рецепторов — это преимущественно пищевые продукты, полученные из растительной пищи, особенно из овощей семейства крестоцветных.Через арилуглеводородные рецепторы лиганды стимулируют иммунную функцию кишечника и гомеостаз кишечника (95). Поскольку лиганды арил-углеводородных рецепторов происходят из кишечной микробиоты, любое нарушение кишечной микробиоты, например, из-за диеты с высоким содержанием жиров, может уменьшить лиганды арил-углеводородных рецепторов. В свою очередь, это может вызвать воспаление кишечника и его проницаемость, а также способствовать развитию метаболического синдрома, который можно улучшить, добавив штамм Lactobacillus (96). Кроме того, уменьшение количества арил-углеводородных рецепторов или лигандов ставит под угрозу поддержание интраэпителиальных лимфоцитов и контроль микробной нагрузки и состава, что приводит к повышенной иммунной активации и повреждению эпителия (95).
Вторичные желчные кислоты и копростанол
Отдельную группу постбиотиков составляют метаболиты холестерина. Описано несколько бактериальных штаммов, выделенных из кишечника или фекалий, для превращения диетического или синтезированного холестерина de novo в копростанол (97, 98), который плохо всасывается в кишечнике человека. Таким образом, уровень холестерина в сыворотке крови хозяина снижается, что снижает риск сердечно-сосудистых заболеваний. С другой стороны, желчные кислоты, синтезированные из холестерина, превращаются микробиотой во вторичные желчные кислоты, обнаруживаемые в различных тканях и в кале.Считается, что вторичные желчные кислоты участвуют в балансе здоровья / болезни (73, 97). Например, они связаны с воспалительным заболеванием кишечника, раком печени и толстой кишки.
N-оксид триметиламина (TMAO)
N-оксид триметиламина — это микробный метаболит, который, как считается, связан с сердечно-сосудистыми и неврологическими расстройствами. Карнитин и холин являются предшественниками ТМАО и в основном содержатся в продуктах животного происхождения (яйца, говядина, свинина), а меньшие количества содержатся в бобах и рыбе (99).Некоторые виды микробов, такие как L- Ruminococcus , связаны с потреблением животных белков и жиров и связаны с уровнями ТМАО (77). В целом, потребление мяса способствует размножению видов Bacteroides, Alistipes, Ruminococcus, Clostridia и Bilophila и умирает Bifidobacterium . Более высокие уровни ТМАО также наблюдаются при потреблении красного мяса, что увеличивает риск сердечно-сосудистых заболеваний и воспалительных заболеваний кишечника (54, 66).У вегетарианцев состав микробиоты кишечника отличается от состава всеядных животных с пониженной способностью вырабатывать триметиламин (ТМА), предшественник ТМАО. Концентрации ТМАО в плазме крови у веганов и лакто-ово-вегетарианцев примерно одинаковы (99, 100).
Снижение уровня ТМАО может быть достигнуто за счет более строгого соблюдения средиземноморской диеты, особенно вегетарианской, богатой фруктами и овощами (77, 100). Повышенное потребление овощей снижает уровень ТМАО за счет уменьшения количества ферментов, ответственных за преобразование ТМА в ТМАО, и за счет ремоделирования микробиоты кишечника.Исследования показали, что у вегетарианцев снижается выработка ТМАО, что снижает их сердечно-сосудистый риск. Чтобы быть объективными, мы должны упомянуть недавнее исследование, оставляющее место для дальнейшего анализа. Трансплантация веганской фекальной микробиоты пациентам с метаболическим синдромом привела к значительным изменениям в составе кишечной микробиоты, но не выявила изменений в продукции ТМАО. Авторы объяснили, что двухнедельное наблюдение было недостаточным для наблюдения изменений в продукции ТМАО (101).
В среднем, двадцать пять процентов метаболитов в плазме у всеядных животных и веганов различаются, что свидетельствует о значительном прямом влиянии диеты на метаболом хозяина. Никакие уникальные бактериальные таксоны не были достоверно связаны с индивидуальными уровнями метаболитов после корректировки для множественных сравнений (102). Эти результаты показывают, что, хотя существует различие между индивидуумами, режимы питания значительно влияют на микробный состав.
Заключение
Текущие исследования показывают, что диета является важным фактором для состава микробиоты кишечника человека, что, в свою очередь, имеет решающее значение для превращения питательных веществ в активные для хозяина постбиотики.Современные знания показывают, что диета на основе растений может быть эффективным способом развития разнообразной экосистемы полезных микробов, поддерживающих общее состояние здоровья. Тем не менее, из-за сложности и индивидуальных различий необходимы дальнейшие исследования, чтобы полностью охарактеризовать взаимодействие между диетой, микробиомом и результатами для здоровья.
Авторские взносы
AT и IB внесли свой вклад в концепцию и написание рукописи, ER, WY, JA, NB и HK внесли свой вклад и критически отредактировали рукопись.Все авторы одобрили окончательную рукопись.
Финансирование
Работа финансировалась ПКРМ и поддержана Грантовым агентством Министерства образования, науки, исследований и спорта Словацкой Республики VEGA 1/0286/18.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Сноски
Список литературы
2.Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C и др. Каталог микробных генов кишечника человека, созданный путем метагеномного секвенирования. Природа. (2010) 464: 59–65. DOI: 10.1038 / nature08821
PubMed Аннотация | CrossRef Полный текст | Google Scholar
3. Уилер Д.А., Сринивасан М., Эгхолм М., Шен Й., Чен Л., Макгуайр А. и др. Полный геном человека путем массового параллельного секвенирования ДНК. Природа. (2008) 452: 872–6. DOI: 10.1038 / nature06884
PubMed Аннотация | CrossRef Полный текст | Google Scholar
6.Roager HM, Licht TR, Poulsen SK, Larsen TM, Bahl MI. Микробные энтеротипы, определяемые соотношением превотеллы и бактероидов, оставались стабильными в течение 6-месячного рандомизированного контролируемого диетического вмешательства с новой нордической диетой. Appl Environ Microbiol. (2014) 80: 1142–9. DOI: 10.1128 / AEM.03549-13
PubMed Аннотация | CrossRef Полный текст | Google Scholar
7. Бык MJ, Plummer NT. Часть 1. Микробиом кишечника человека в условиях здоровья и болезней. Integr Med Clin J. (2014) 13: 17–22.
PubMed Аннотация | Google Scholar
10. Ларсен Н., Фогенсен Ф.К., ван ден Берг Ф.В., Нильсен Д.С., Андреасен А.С., Педерсен Б.К. и др. Микробиота кишечника взрослых людей с диабетом 2 типа отличается от взрослых людей, не страдающих диабетом. PLoS ONE. (2010) 5: e9085. DOI: 10.1371 / journal.pone.0009085
PubMed Аннотация | CrossRef Полный текст | Google Scholar
14. Унгер М.М., Шпигель Дж., Диллманн К.Ю., Грундманн Д., Филипп Х., Бюрманн Дж. И др. Короткоцепочечные жирные кислоты и микробиота кишечника различаются между пациентами с болезнью Паркинсона и контрольной группой того же возраста. Паркинсонизм, связанный с расстройством. (2016) 32: 66–72. DOI: 10.1016 / j.parkreldis.2016.08.019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
17. Томова А., Хусарова В., Лакатосова С., Бакос Дж., Влкова Б., Бабинская К. и др. Микробиота желудочно-кишечного тракта у детей с аутизмом в Словакии. Physiol Behav. (2015) 138: 179–87. DOI: 10.1016 / j.physbeh.2014.10.033
PubMed Аннотация | CrossRef Полный текст | Google Scholar
18. Селбер-Хнатив С., Рукундо Б., Ахмади М., Акуби Х., Аль-Бизри Х., Алиу А.Ф. и др.Микробиота кишечника человека: к экологии болезней. Front Microbiol. (2017) 8: 1265. DOI: 10.3389 / fmicb.2017.01265
PubMed Аннотация | CrossRef Полный текст | Google Scholar
20. Ву Г.Д., Чен Дж., Хоффманн С., Биттингер К., Чен Ю.Й., Кейлбо С.А. и др. Связывание долгосрочных диетических моделей с кишечными микробными энтеротипами. Наука. (2011) 334: 105–8. DOI: 10.1126 / science.1208344
PubMed Аннотация | CrossRef Полный текст | Google Scholar
22.Ян В., Чжэн Дж., Вэнь Ц., Цзи Ц., Чжан Д., Чен Й и др. Эффективность анализа фекалий в качестве индикатора кишечника при исследовании микробиоты кишечника цыплят. bioRxiv. 2018: 313577. DOI: 10.1101 / 313577
CrossRef Полный текст
26. Wong MW, Yi CH, Liu TT, Lei WY, Hung JS, Lin CL, et al. Влияние веганской диеты на микробиоту кишечника: обновленная информация о клинических последствиях. Tzu Chi Med J. (2018) 30: 200–3. DOI: 10.4103 / tcmj.tcmj_21_18
PubMed Аннотация | CrossRef Полный текст | Google Scholar
27.Клименко Н.С., Тяхт А.В., Попенко А.С., Васильев А.С., Алтухов И.А., Ищенко Д.С. и др. Реакция микробиома на неконтролируемое краткосрочное диетическое вмешательство в рамках проекта гражданской науки. Питательные вещества. (2018) 10: E576. DOI: 10.3390 / nu10050576
PubMed Аннотация | CrossRef Полный текст | Google Scholar
28. Мартинес И., Латтимер Дж. М., Хубах К. Л., Кейс Дж. А., Янг Дж., Вебер К. Г. и др. Состав микробиома кишечника связан с иммунологическими улучшениями, вызванными цельным зерном. ISME J. (2013) 7: 269–80. DOI: 10.1038 / ismej.2012.104
PubMed Аннотация | CrossRef Полный текст | Google Scholar
29. Циммер Дж., Ланге Б., Фрик Дж. С., Зауэр Х., Циммерманн К., Швирц А. и др. Вегетарианская или вегетарианская диета существенно изменяет фекальную микробиоту толстой кишки человека. Eur J Clin Nutr. (2012) 66: 53–60. DOI: 10.1038 / ejcn.2011.141
PubMed Аннотация | CrossRef Полный текст | Google Scholar
30. Скотт К.П., Дункан С.Х., Флинт Х.Дж.Пищевые волокна и микробиота кишечника. Nutr Bull. (2008). 33: 201–211. DOI: 10.1111 / j.1467-3010.2008.00706.x
CrossRef Полный текст | Google Scholar
31. Вердам Ф. Дж., Фуэнтес С., де Йонге С., Зоетендал Э. Г., Эрбиль Р., Греве Дж. В. и др. Состав кишечной микробиоты человека связан с местным и системным воспалением при ожирении. Obes Silver Spring Md. (2013) 21: E607–615. DOI: 10.1002 / oby.20466
PubMed Аннотация | CrossRef Полный текст | Google Scholar
32.Орлич MJ, Fraser GE. Вегетарианские диеты в исследовании Adventist Health Study 2: обзор первых опубликованных результатов1234. Am J Clin Nutr. (2014) 100: 353S-8S. DOI: 10.3945 / ajcn.113.071233
PubMed Аннотация | CrossRef Полный текст | Google Scholar
33. Menni C, Lin C, Cecelja M, Mangino M, Matey-Hernandez ML, Keehn L, et al. Разнообразие кишечных микробов связано с более низкой жесткостью артерий у женщин. Eur Heart J. (2018) 39: 2390–7. DOI: 10.1093 / eurheartj / ehy226
PubMed Аннотация | CrossRef Полный текст | Google Scholar
34.Риццони Д., Портери Э., Гуэлфи Д., Муйесан М.Л., Валентини Ю., Чимино А. и др. Структурные изменения подкожных мелких артерий нормотензивных и гипертензивных пациентов с инсулинозависимым сахарным диабетом. Тираж. (2001) 103: 1238–44. DOI: 10.1161 / 01.CIR.103.9.1238
PubMed Аннотация | CrossRef Полный текст | Google Scholar
35. Losasso C, Eckert EM, Mastrorilli E, Villiger J, Mancin M, Patuzzi I, et al. Оценка влияния западных диетических стилей веганской, вегетарианской и всеядной диеты на микробиоту кишечника человека: перекрестное исследование. Front Microbiol. (2018) 9: 317. DOI: 10.3389 / fmicb.2018.00317
PubMed Аннотация | CrossRef Полный текст | Google Scholar
36. do Rosario VA, Fernandes R, Trindade EB. Вегетарианские диеты и микробиота кишечника: важные сдвиги в маркерах обмена веществ и сердечно-сосудистых заболеваний. Nutr Ред. (2016) 74: 444–54. DOI: 10.1093 / Nutrit / nuw012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
38. Бамбергер С., Россмайер А., Лехнер К., Ву Л., Вальдманн Е., Фишер С. и др.Обогащенная грецкими орехами диета влияет на микробиом кишечника у здоровых людей европеоидной расы: рандомизированное контролируемое исследование. Питательные вещества. (2018) 10. DOI: 10.3390 / nu10020244
PubMed Аннотация | CrossRef Полный текст | Google Scholar
39. Джайн А., Ли XH, Чен В.Н. Сходства и различия в составе микробиома кишечника коррелируют с диетами взрослых индийцев и китайцев. AMB Express. (2018) 8: 104. DOI: 10.1186 / s13568-018-0632-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
40.Де Филиппо С., Кавальери Д., Ди Паола М., Рамазотти М., Пулле Дж. Б., Массарт С. и др. Влияние диеты на формирование микробиоты кишечника выявлено в сравнительном исследовании у детей из Европы и сельских районов Африки. Proc Natl Acad Sci USA. (2010) 107: 14691–6. DOI: 10.1073 / pnas.1005963107
PubMed Аннотация | CrossRef Полный текст | Google Scholar
41. Мартинес И., Ким Дж., Даффи П. Р., Шлегель В. Л., Вальтер Дж. Устойчивые крахмалы 2 и 4 типов по-разному влияют на состав фекальной микробиоты у людей. PloS ONE. (2010) 5: e15046. DOI: 10.1371 / journal.pone.0015046
PubMed Аннотация | CrossRef Полный текст | Google Scholar
42. Лей Р. Э., Бэкхед Ф., Тернбо П., Лозупоне, Калифорния, Найт Р. Д., Гордон Дж. И.. Ожирение изменяет микробную экологию кишечника. Proc Natl Acad Sci USA. (2005) 102: 11070–5. DOI: 10.1073 / pnas.0504978102
PubMed Аннотация | CrossRef Полный текст | Google Scholar
43. Лин А., Бик Е.М., Костелло Е.К., Детлефсен Л., Хак Р., Релман Д.А. и др.Отчетливое разнообразие и состав микробиома дистальных отделов кишечника у здоровых детей из Бангладеш и США. PloS ONE. (2013) 8: e53838. DOI: 10.1371 / journal.pone.0053838
PubMed Аннотация | CrossRef Полный текст | Google Scholar
44. Швирц А., Тарас Д., Шефер К., Бейер С., Бос Н.А., Донус С. и др. Микробиота и SCFA у худых и здоровых людей с избыточным весом. Obes Silver Spring Md. (2010) 18: 190–5. DOI: 10.1038 / oby.2009.167
PubMed Аннотация | CrossRef Полный текст | Google Scholar
45.Андох А., Нишида А., Такахаши К., Инатоми О, Имаеда Х., Бамба С. и др. Сравнение микробного сообщества кишечника у людей с ожирением и худых с использованием секвенирования гена 16S в популяции Японии. J Clin Biochem Nutr. (2016) 59: 65–70. DOI: 10.3164 / jcbn.15-152
PubMed Аннотация | CrossRef Полный текст | Google Scholar
46. Клэссон М.Дж., Джеффри И.Б., Конде С., Пауэр С.Е., О’Коннор Е.М., Кьюсак С. и др. Состав кишечной микробиоты у пожилых людей коррелирует с диетой и здоровьем. Природа. (2012) 488: 178–84. DOI: 10.1038 / природа11319
PubMed Аннотация | CrossRef Полный текст | Google Scholar
47. Руенгсомвонг С., Ла-Онгкхам О, Цзян Дж., Ванниссорн Б., Накаяма Дж., Нитисинпрасерт С. Микробное сообщество здоровых тайских вегетарианцев и невегетарианцев, их внутренняя микробиота кишечника и риск патогенов. J Microbiol Biotechnol. (2016) 26: 1723–35. DOI: 10.4014 / jmb.1603.03057
PubMed Аннотация | CrossRef Полный текст | Google Scholar
49.Матияшич Б. Б., Обермайер Т., Липоглавшек Л., Грабнар И., Авгуштин Г., Рогель И. Связь диетического типа с фекальной микробиотой у вегетарианцев и всеядных животных в Словении. Eur J Nutr. (2014) 53: 1051–64. DOI: 10.1007 / s00394-013-0607-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
50. Ковачева-Датчари П., Нильссон А., Акрами Р., Ли Ю.С., Де Ваддер Ф., Арора Т. и др. Улучшение метаболизма глюкозы, вызванное пищевыми волокнами, связано с увеличением количества превотеллы. Cell Metab. (2015) 22: 971–82. DOI: 10.1016 / j.cmet.2015.10.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
51. Дэвид Л.А., Морис К.Ф., Кармоди Р.Н., Гутенберг Д.Б., Баттон Дж. Э., Вулф Б. Э. и др. Диета быстро и воспроизводимо изменяет микробиом кишечника человека. Природа. (2014) 505: 559–63. DOI: 10.1038 / природа12820
PubMed Аннотация | CrossRef Полный текст | Google Scholar
52. Whisner CM, Maldonado J, Dente B, Krajmalnik-Brown R, Bruening M.Диета, физическая активность и экранное время, но не индекс массы тела, связаны с микробиомом кишечника разнообразной когорты студентов колледжей, проживающих в университетских корпусах: перекрестное исследование. BMC Microbiol. (2018) 18: 210. DOI: 10.1186 / s12866-018-1362-x
CrossRef Полный текст | Google Scholar
53. Исии К., Наканиси Ю., Мураками С., Нозу Р., Уэно М., Хиоки К. и др. Метабологеномный подход позволяет выявить изменения в кишечной среде мышей, получавших американскую диету. Int J Mol Sci. (2018) 19: E4079. DOI: 10.3390 / ijms1
79
PubMed Аннотация | CrossRef Полный текст | Google Scholar
54. Сингх Р.К., Чанг Х.В., Ян Д., Ли К.М., Укмак Д., Вонг К. и др. Влияние диеты на микробиом кишечника и последствия для здоровья человека. J Transl Med. (2017) 15:73. DOI: 10.1186 / s12967-017-1175-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
56. Чжан С., Бьоркман А., Цай К., Лю Г., Ван С., Ли И и др.Влияние трехмесячной вегетарианской диеты на микробиоту кишечника и иммунный репертуар. Front Immunol. (2018) 9: 908. DOI: 10.3389 / fimmu.2018.00908
PubMed Аннотация | CrossRef Полный текст | Google Scholar
59. Уилан К., Джадд П.А., Приди В.Р., Симмеринг Р., Янн А., Тейлор М.А. Фруктоолигосахариды и клетчатка частично предотвращают изменения фекальной микробиоты и концентрации короткоцепочечных жирных кислот, вызываемые стандартной энтеральной формулой у здоровых людей. J Nutr. (2005) 135: 1896–902. DOI: 10.1093 / JN / 135.8.1896
PubMed Аннотация | CrossRef Полный текст | Google Scholar
60. Ten Bruggencate SJ, Bovee-Oudenhoven IM, Lettink-Wissink ML, Katan MB, van der Meer R. Диетические фруктоолигосахариды влияют на барьерную функцию кишечника у здоровых мужчин. J Nutr. (2006) 136: 70–4. DOI: 10.1093 / jn / 136.1.70
PubMed Аннотация | CrossRef Полный текст | Google Scholar
61. Чен Т., Лонг В., Чжан С., Лю С., Чжао Л., Хамакер Б.Р.Способность использовать клетчатку варьируется в кишечной микробиоте с преобладанием Prevotella и Bacteroides. Научный доклад (2017) 7: 2594. DOI: 10.1038 / s41598-017-02995-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
62. Fehlbaum S, Prudence K, Kieboom J, Heerikhuisen M, van den Broek T., Schuren FHJ, et al. In vitro ферментация выбранных пребиотиков и их влияние на состав и активность микробиоты кишечника взрослых. Int J Mol Sci. (2018) 19:10.DOI: 10.3390 / ijms1
97
PubMed Аннотация | CrossRef Полный текст | Google Scholar
63. Уилсон Б., Уилан К. Пребиотические фруктаны инулинового типа и галактоолигосахариды: определение, специфичность, функция и применение при желудочно-кишечных расстройствах. J Gastroenterol Hepatol. (2017) 32: 64–8. DOI: 10.1111 / jgh.13700
PubMed Аннотация | CrossRef Полный текст | Google Scholar
64. Hentges DJ, Майер Б.Р., Бертон Г.К., Флинн М.А., Цутакава Р.К. Влияние диеты с высоким содержанием говядины на фекальную бактериальную флору человека. Cancer Res. (1977) 37: 568–71.
PubMed Аннотация | Google Scholar
65. Sheflin AM, Melby CL, Carbonero F, Weir TL. Связь режима питания с микробным составом и функцией кишечника. Кишечные микробы. (2016) 8: 113–29. DOI: 10.1080 / 194
.2016.1270809
PubMed Аннотация | CrossRef Полный текст | Google Scholar
67. Коэльо О.Г., Кандидо Ф.Г., Альфенас РКГ. Диетический жир и микробиота кишечника: механизмы, участвующие в контроле ожирения. Crit Rev Food Sci Nutr. (2018) 2018: 1–9. DOI: 10.1080 / 10408398.2018.1481821
CrossRef Полный текст | Google Scholar
69. Сунь Х., Чен И, Ченг М., Чжан Х, Чжэн Х, Чжан З. Модулирующее действие полифенолов из зеленого чая, чая улун и черного чая на микробиоту кишечника человека in vitro. J Food Sci Technol. (2018) 55: 399–407. DOI: 10.1007 / s13197-017-2951-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
70.Клемашевич C, Wu C, Howsmon D, Alaniz RC, Lee K, Jayaraman A. Рациональная идентификация постбиотиков, полученных из диеты, для улучшения функции кишечной микробиоты. Curr Opin Biotechnol. (2014) 26: 85–90. DOI: 10.1016 / j.copbio.2013.10.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
72. Сравните D, Coccoli P, Rocco A, Nardone OM, De Maria S, Cartenì M и др. Ось кишечник – печень: влияние микробиоты кишечника на неалкогольную жировую болезнь печени. Nutr Metab Cardiovasc Dis. (2012) 22: 471–6. DOI: 10.1016 / j.numecd.2012.02.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
74. Агилар-Тоала Дж. Э., Гарсия-Варела Р., Гарсия Х. С. и др. Постбиотики: развивающийся термин в области функциональных продуктов питания. Trends Food Sci Technol. (2018) 75: 105–14. DOI: 10.1016 / j.tifs.2018.03.009
CrossRef Полный текст | Google Scholar
75. Моррисон Д. Д., Престон Т. Образование короткоцепочечных жирных кислот кишечной микробиотой и их влияние на метаболизм человека. Кишечные микробы. (2016) 7: 189–200. DOI: 10.1080 / 194
.2015.1134082
PubMed Аннотация | CrossRef Полный текст | Google Scholar
76. Cani PD, Bibiloni R, Knauf C, et al. Изменения микробиоты кишечника контролируют воспаление, вызванное метаболической эндотоксемией, при ожирении и диабете, вызванном диетой с высоким содержанием жиров. Диабет. (2008) 57: 1470–81. DOI: 10.2337 / db07-1403
PubMed Аннотация | CrossRef Полный текст | Google Scholar
77. Де Филиппис Ф., Пеллегрини Н., Ваннини Л., Джеффри И.Б., Ла Сториа А., Лаги Л. и др.Строгое соблюдение средиземноморской диеты благотворно влияет на микробиоту кишечника и связанный с ней метаболом. Gut. (2016) 65: 1812–21. DOI: 10.1136 / gutjnl-2015-309957
PubMed Аннотация | CrossRef Полный текст | Google Scholar
78. Ко А., Де Ваддер Ф, Ковачева-Датчари П., Бекхед Ф. От пищевых волокон к физиологии хозяина: короткоцепочечные жирные кислоты как ключевые бактериальные метаболиты. Ячейка. (2016) 165: 1332–45. DOI: 10.1016 / j.cell.2016.05.041
PubMed Аннотация | CrossRef Полный текст | Google Scholar
80.Рейнес Б., Палоу М., Родригес А.М., Палоу А. Регулирование адаптивного термогенеза и потемнения пребиотиками и постбиотиками. Front Physiol. (2019) 9: 1908. DOI: 10.3389 / fphys.2018.01908
PubMed Аннотация | CrossRef Полный текст | Google Scholar
82. Velázquez OC, Lederer HM, Rombeau JL. Бутират и колоноцит. Производство, абсорбция, метаболизм и терапевтические последствия. Adv Exp Med Biol. (1997) 427: 123–34. DOI: 10.1007 / 978-1-4615-5967-2_14
PubMed Аннотация | CrossRef Полный текст | Google Scholar
85.Оно К., Ли Л., Такамура Ю., Йошиике Ю., Чжу Л., Хань Ф. и др. Фенольные соединения предотвращают олигомеризацию амилоидного β-белка и синаптическую дисфункцию за счет сайт-специфического связывания. J Biol Chem. (2012) 287: 14631–43. DOI: 10.1074 / jbc.M111.325456
PubMed Аннотация | CrossRef Полный текст | Google Scholar
86. Хоссен М.С., Али М.И., Джахурул М.Х., Абдель-Дайм М.М., Ган С.Х., Халил М.И. Полезная роль полифенолов меда против некоторых дегенеративных заболеваний человека: обзор. Pharmacol Rep. (2017) 69: 1194–205. DOI: 10.1016 / j.pharep.2017.07.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
87. Ландете Дж. М., Аркес Дж., Медина М., Гая П., де Лас Ривас Б., Муньос Р. Биоактивация фитоэстрогенов: кишечные бактерии и здоровье. Crit Rev Food Sci Nutr. (2016) 56: 1826–43. DOI: 10.1080 / 10408398.2013.789823
PubMed Аннотация | CrossRef Полный текст | Google Scholar
88. Томас-Барберан Ф.А., Гонсалес-Сарриас А., Гарсия-Вильяльба Р., Нуньес-Санчес М.А., Сельма М.В., Гарсия-Конеса М.Т. и др.Уролитины, спасение «старых» метаболитов для понимания «новой» концепции: метаботипы как связующее звено между фенольным метаболизмом, дисбиозом микробиоты и состоянием здоровья хозяина. Mol Nutr Food Res. (2017) 61: 1. DOI: 10.1002 / mnfr.201500901
PubMed Аннотация | CrossRef Полный текст | Google Scholar
90. Cardona F, Andrés-Lacueva C, Tulipani S, Tinahones FJ, Queipo-Ortuño MI. Польза полифенолов для микробиоты кишечника и их влияние на здоровье человека. J Nutr Biochem. (2013) 24: 1415–22. DOI: 10.1016 / j.jnutbio.2013.05.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
91. Ozdal T, Sela DA, Xiao J, Boyacioglu D, Chen F, Capanoglu E. Взаимодействие между полифенолами и кишечной микробиотой и влияние на биодоступность. Питательные вещества. (2016) 8: 2. DOI: 10.3390 / nu8020078
PubMed Аннотация | CrossRef Полный текст | Google Scholar
92. De Filippis F, Pellegrini N, Laghi L, Gobbetti M, Ercolini D.Необычные ассоциации подрода фекальных Prevotella и Bacteroides с определенными диетическими моделями. Микробиом. (2016) 4. DOI: 10.1186 / s40168-016-0202-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
93. ЛеБлан Дж. Г., Милани С., де Джорджи Г. С., Сесма Ф, ван Синдерен Д., Вентура М. Бактерии как поставщики витаминов для своего хозяина: взгляд на микробиоту кишечника. Curr Opin Biotechnol. (2013) 24: 160–8. DOI: 10.1016 / j.copbio.2012.08.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
94.Тиан С., Лю Х, Лей П., Чжан Х, Шань Ю. Микробиота: медиатор для преобразования предшественников глюкозинолатов в крестоцветных овощах в активные изотиоцианаты. J Sci Food Agric. (2018) 98: 1255–60. DOI: 10.1002 / jsfa.8654
PubMed Аннотация | CrossRef Полный текст | Google Scholar
95. Li Y, Innocentin S, Withers DR, Roberts NA, Gallagher AR, Grigorieva EF, et al. Экзогенные стимулы поддерживают интраэпителиальные лимфоциты за счет активации рецепторов арилуглеводородов. Ячейка. (2011) 147: 629–40. DOI: 10.1016 / j.cell.2011.09.025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
96. Нативидад Дж. М., Ламас Б., Фам Х. П., Мишель М. Л., Рейнто Д., Бридонно С. и др. Bilophila wadsworthia усугубляет метаболические дисфункции у мышей, вызванные диетой с высоким содержанием жиров. Nat Commun. (2018) 9: 2802. DOI: 10.1038 / s41467-018-05249-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
98. Horáčková Š, Plocková M, Demnerová K.Важность систем микробной защиты для солей желчных кислот и механизмов снижения уровня холестерина в сыворотке. Biotechnol Adv. (2018) 36: 682–90. DOI: 10.1016 / j.biotechadv.2017.12.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
99. Жанейро М. Х., Рамирес М. Дж., Милагро Ф. И., Мартинес Дж. А., Солас М. Влияние N-оксида триметиламина (ТМАО) на заболевание: потенциальный биомаркер или новая терапевтическая мишень. Питательные вещества. (2018) 10:10. DOI: 10.3390 / nu10101398
CrossRef Полный текст | Google Scholar
101.Smits LP, Kootte RS, Levin E, Prodan A, Fuentes S, Zoetendal EG, et al. Влияние трансплантации веганской фекальной микробиоты на продукцию триметиламин-N-оксида карнитина и холина и воспаление сосудов у пациентов с метаболическим синдромом. J Am Heart Assoc. (2018) 7: 7. DOI: 10.1161 / JAHA.117.008342
PubMed Аннотация | CrossRef Полный текст | Google Scholar
102. Wu GD, Compher C, Chen EZ, Smith SA, Shah RD, Bittinger K, et al. Сравнительная метаболомика у веганов и всеядных животных выявляет ограничения на производство метаболитов кишечной микробиоты, зависящее от диеты. Gut. (2016) 65: 63–72. DOI: 10.1136 / gutjnl-2014-308209
PubMed Аннотация | CrossRef Полный текст | Google Scholar
103. Хаяси Х., Сакамото М., Бенно Ю. Микробное разнообразие фекалий у строгих вегетарианцев, определенное молекулярным анализом и культивированием. Microbiol Immunol. (2002) 46: 819–31. DOI: 10.1111 / j.1348-0421.2002.tb02769.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Антибактериальная и антибактериальная активность ауранофина против Bacteroides fragilis
АбдельХалек А., Абуталеб Н.С., Эльмагармид К.А., Селим М.Н. (2018) Использование ауранофина в качестве средства деколонизации кишечника для устойчивых к ванкомицину энтерокицин.Sci Rep 8: 8353. https://doi.org/10.1038/s41598-018-26674-0
CAS Статья PubMed PubMed Central Google Scholar
Abouelhassan Y et al (2017) Нитроксолин: средство широкого спектра действия для уничтожения биопленок против патогенных бактерий. Int J Antimicrob Agents 49: 247–251. https://doi.org/10.1016/j.ijantimicag.2016.10.017
CAS Статья PubMed Google Scholar
Алав И., Саттон Дж. М., Рахман К. М. (2018) Роль насосов для оттока бактерий в формировании биопленки.Журнал Antimicrob Chemother 73: 2003–2020. https://doi.org/10.1093/jac/dky042
CAS Статья PubMed Google Scholar
Boleij A et al (2015) Ген токсина Bacteroides fragilis преобладает в слизистой оболочке толстой кишки пациентов с колоректальным раком. Clin Infect Dis 60: 208–215. https://doi.org/10.1093/cid/ciu787
CAS Статья PubMed Google Scholar
Bundgaard-Nielsen C, Baandrup UT, Nielsen LP, Sorensen S (2019) Присутствие бактерий варьируется в зависимости от колоректальной аденокарциномы, предшественников поражений и доброкачественной ткани.BMC Рак 19: 399. https://doi.org/10.1186/s12885-019-5571-y
Артикул PubMed PubMed Central Google Scholar
Chen H, Wubbolts RW, Haagsman HP, Veldhuizen EJA (2018) Ингибирование и искоренение биопленок Pseudomonas aeruginosa пептидами защиты хозяина. Sci Rep 8: 10446. https://doi.org/10.1038/s41598-018-28842-8
CAS Статья PubMed PubMed Central Google Scholar
Институт клинических и лабораторных стандартов (CLSI) (2018) Методы тестирования антимикробной чувствительности анаэробных бактерий, 9-е изд.CLSI, Уэйн, стр. M11
Google Scholar
Costa OYA, Raaijmakers JM, Kuramae EE (2018) Микробные внеклеточные полимерные вещества: экологическая функция и влияние на агрегацию почвы. Front Microbiol 9: 1636. https://doi.org/10.3389/fmicb.2018.01636
Артикул PubMed PubMed Central Google Scholar
Debnath A et al (2012). Высокопроизводительный скрининг лекарств для Entamoeba histolytica позволяет идентифицировать новое отведение и цель.Nat Med 18: 956–960. https://doi.org/10.1038/nm.2758
CAS Статья PubMed PubMed Central Google Scholar
de Vos WM (2015) Микробные биопленки и микробиом кишечника человека. NPJ Biofilms Microbiomes 1: 15005. https://doi.org/10.1038/npjbiofilms.2015.5
CAS Статья PubMed PubMed Central Google Scholar
Donelli G, Vuotto C, Cardines R, Mastrantonio P (2012) Растущие на биопленке кишечные анаэробные бактерии.FEMS Immunol Med Microbiol 65: 318–325. https://doi.org/10.1111/j.1574-695X.2012.00962.x
CAS Статья PubMed Google Scholar
Felipe V et al (2019) Хитозан нарушает образование биопленок и способствует уничтожению биопленок у видов Staphylococcus, выделенных от мастита крупного рогатого скота. Int J Biol Macromol 126: 60–67. https://doi.org/10.1016/j.ijbiomac.2018.12.159
CAS Статья PubMed Google Scholar
Flemming HC, Neu TR, Wozniak DJ (2007) Матрица EPS: «дом клеток биопленки».J Bacteriol 189: 7945–7947. https://doi.org/10.1128/JB.00858-07
CAS Статья PubMed PubMed Central Google Scholar
Goel S, Mishra P (2018) Тимохинон подавляет образование биопленок и обладает избирательной антибактериальной активностью за счет образования ROS. Appl Microbiol Biotechnol 102: 1955–1967. https://doi.org/10.1007/s00253-018-8736-8
CAS Статья PubMed Google Scholar
Harbut MB et al (2015) Ауранофин оказывает бактерицидное действие широкого спектра, воздействуя на тиоловый окислительно-восстановительный гомеостаз.Proc Natl Acad Sci USA 112: 4453–4458. https://doi.org/10.1073/pnas.1504022112
CAS Статья PubMed Google Scholar
Holmgren A, Lu J (2010) Тиоредоксин и тиоредоксинредуктаза: текущие исследования с особым вниманием к болезням человека. Biochem Biophys Res Commun. 396: 120–124. https://doi.org/10.1016/j.bbrc.2010.03.083
CAS Статья PubMed Google Scholar
Hwang-Bo H et al (2017) Ауранофин, ингибитор тиоредоксинредуктазы, индуцирует апоптоз в клетках гепатоцеллюлярной карциномы Hep3B путем генерации активных форм кислорода.Gen Physiol Biophys 36: 117–128. https://doi.org/10.4149/gpb_2016043
CAS Статья PubMed Google Scholar
Joo HS, Otto M (2012) Молекулярная основа образования биопленок in vivo бактериальными патогенами. Chem Biol 19: 1503–1513. https://doi.org/10.1016/j.chembiol.2012.10.022
CAS Статья PubMed PubMed Central Google Scholar
Като Н. и др. (2000) Новый подтип гена токсина металлопротеиназы и встречаемость трех подтипов bft среди изолятов Bacteroides fragilis в Японии.FEMS Microbiol Lett 182: 171–176. https://doi.org/10.1111/j.1574-6968.2000.tb08892.x
CAS Статья PubMed Google Scholar
Kim HR, Rhee KJ, Eom YB (2019) Антибиопленочные и антимикробные эффекты зерумбона против Bacteroides fragilis . Анаэроб 57: 99–106. https://doi.org/10.1016/j.anaerobe.2019.04.001
CAS Статья PubMed Google Scholar
Ковальский Р.П., Ятс К.А., Романовски Э.Г., Каренчак Л.М., Мах Ф.С., Гордон Ю.Дж. (2005) Руководство офтальмолога для понимания чувствительности к антибиотикам и данных о минимальной ингибирующей концентрации.Офтальмология 112: 1987. https://doi.org/10.1016/j.ophtha.2005.06.025
Артикул PubMed Google Scholar
Mendes RF et al (2017) Эфирное масло из плодов бразильской пряности Xylopia sericea A. St.-Hil. проявляет выраженную антибактериальную и антиоксидантную активность in vitro. J Pharm Pharmacol 69: 341–348. https://doi.org/10.1111/jphp.12698
CAS Статья PubMed Google Scholar
Молина-Мансо Д. и др. (2013) Чувствительность стафилококков к антибиотикам в биопленках, выделенных из ортопедических инфекций, in vitro.Int J Antimicrob Agents 41: 521–523. https://doi.org/10.1016/j.ijantimicag.2013.02.018
CAS Статья PubMed Google Scholar
Myers LL, Shoop DS, Collins JE, Bradbury WC (1989) Диарейное заболевание, вызванное энтеротоксигенным Bacteroides fragilis у новорожденных кроликов. J Clin Microbiol 27: 2025–2030
CAS Статья Google Scholar
Park SH, Lee JH, Berek JS, Hu MC (2014) Ауранофин проявляет противораковую активность против клеток рака яичников посредством активации FOXO3 независимо от p53.Int J Oncol 45: 1691–1698. https://doi.org/10.3892/ijo.2014.2579
CAS Статья PubMed PubMed Central Google Scholar
Pierce JV, Bernstein HD (2016) Геномное разнообразие энтеротоксигенных штаммов Bacteroides fragilis . PLoS ONE 11: e0158171. https://doi.org/10.1371/journal.pone.0158171
CAS Статья PubMed PubMed Central Google Scholar
Пул К., Срикумар Р. (2001) Отток нескольких лекарственных препаратов в Pseudomonas aeruginosa : компоненты, механизмы и клиническое значение.Curr Top Med Chem 1: 59–71
CAS Статья Google Scholar
Pumbwe L, Ueda O, Yoshimura F, Chang A, Smith RL, Wexler HM (2006) Bacteroides fragilis Системы оттока BmeABC аддитивно придают внутреннюю устойчивость к противомикробным препаратам. J Antimicrob Chemother 58: 37–46. https://doi.org/10.1093/jac/dkl202
CAS Статья PubMed Google Scholar
Pumbwe L, Skilbeck CA, Wexler HM (2008) Наличие систем контроля кворума, связанных с множественной лекарственной устойчивостью и образованием биопленок у Bacteroides fragilis .Microb Ecol 56: 412–419. https://doi.org/10.1007/s00248-007-9358-3
CAS Статья PubMed Google Scholar
Reichhardt C, Parsek MR (2019) Конфокальная лазерная сканирующая микроскопия для анализа архитектуры биопленок Pseudomonas aeruginosa и локализации матрикса. Front Microbiol 10: 677. https://doi.org/10.3389/fmicb.2019.00677
Артикул PubMed PubMed Central Google Scholar
Rhee KJ et al (2009) Индукция стойкого колита энтеротоксигенным комменсалом человека Bacteroides fragilis у мышей C57BL / 6 дикого типа.Заражение иммунной 77: 1708–1718. https://doi.org/10.1128/IAI.00814-08
CAS Статья PubMed PubMed Central Google Scholar
Rocha ER, Tzianabos AO, Smith CJ (2007) Тиоредоксинредуктаза необходима для тиол-дисульфидного окислительно-восстановительного контроля и выживания анаэроба при окислительном стрессе Bacteroides fragilis . J Bacteriol 189: 8015–8023. https://doi.org/10.1128/JB.00714-07
CAS Статья PubMed PubMed Central Google Scholar
Roder C, Thomson MJ (2015) Ауранофин: перепрофилирование старого препарата для золотого нового века.Исследования и разработки в области лекарственных средств 15: 13–20. https://doi.org/10.1007/s40268-015-0083-y
CAS Статья Google Scholar
Rosenblatt JE, Stewart PR (1975) Метод анаэробного культивирования в мешках. J Clin Microbiol 1: 527–530
CAS Статья Google Scholar
Schwarz S et al (2010) От редакции: оценка чувствительности к противомикробным препаратам бактерий, полученных от животных.Журнал Antimicrob Chemother 65: 601–604. https://doi.org/10.1093/jac/dkq037
CAS Статья PubMed Google Scholar
Смит С.Г., Махон В., Ламберт М.А., Фаган Р.П. (2007) Молекулярный швейцарский армейский нож: структура, функция и выражение OmpA. FEMS Microbiol Lett 273: 1–11. https://doi.org/10.1111/j.1574-6968.2007.00778.x
CAS Статья PubMed Google Scholar
Swidsinski A, Weber J, Loening-Baucke V, Hale LP, Lochs H (2005) Пространственная организация и состав флоры слизистой оболочки у пациентов с воспалительным заболеванием кишечника.J Clin Microbiol 43: 3380–3389. https://doi.org/10.1128/JCM.43.7.3380-3389.2005
Артикул PubMed PubMed Central Google Scholar
Thangamani S et al (2016a) Антибактериальная активность и механизм действия ауранофина против бактериальных патогенов с множественной лекарственной устойчивостью. Sci Rep 6: 22571. https://doi.org/10.1038/srep22571
CAS Статья PubMed PubMed Central Google Scholar
Thangamani S, Mohammad H, Abushahba MF, Sobreira TJ, Seleem MN (2016b) Использование ауранофина для лечения кожных стафилококковых инфекций.Int J Antimicrob Agents 47: 195–201. https://doi.org/10.1016/j.ijantimicag.2015.12.016
CAS Статья PubMed PubMed Central Google Scholar
Wexler HM (2007) Bacteroides: хорошие, плохие и мелкие детали. Clin Microbiol Rev 20: 593–621. https://doi.org/10.1128/CMR.00008-07
CAS Статья PubMed PubMed Central Google Scholar
Wexler HM, Tenorio E, Pumbwe L (2009) Характеристики Bacteroides fragilis , лишенного основного белка внешней мембраны, OmpA.Микробиология 155: 2694–2706. https://doi.org/10.1099/mic.0.025858-0
CAS Статья PubMed Google Scholar
Woo SG et al (2017) Эффективность антибиотикопленочных и антивирулентных свойств дигидроцеластрола и дигидроцеластрилдиацетата в борьбе с метициллин-устойчивым Staphylococcus aureus . Arch Microbiol 199: 1151–1163. https://doi.org/10.1007/s00203-017-1386-x
CAS Статья PubMed Google Scholar
Некротический фасциит — NORD (Национальная организация по редким заболеваниям)
NF вызывается бактерией (мономикробный NF) или несколькими бактериями (полимикробный NF), поражающими ткань непосредственно под кожей (подкожная клетчатка).Бактерии или бактерии попадают в организм либо через внешнюю травму (места хирургического вмешательства, порез, царапину, синяк, фурункул или любую небольшую травму), либо через прямое распространение от проколотого / перфорированного внутреннего органа (особенно толстой кишки, прямой кишки или анус) или половой орган. При заражении бактерии или бактерии распространяются через фасцию, производя эндотоксины (токсины, выделяемые, когда бактерии умирают, распадаются или лизируются) и экзотоксины (токсины, выделяемые бактериями в виде отходов), которые ограничивают кровоснабжение тканей (ткани ишемия), переваривание клеток ферментами, приводящее к поражению, состоящему из гноя и жидких остатков мертвой ткани, и часто системное заболевание (заболевание любой основной системы органов или любое состояние, которое в конечном итоге влияет на все тело).Поскольку кровоснабжение этих тканей нарушается, ни антибиотики, ни собственные механизмы организма для борьбы с инфекцией не могут достичь этих тканей. Таким образом, лечение требует хирургической обработки раны (хирургического удаления мертвых и инфицированных тканей).
Учитывая распространенность всех бактерий, вызывающих ЯФ, важно следить за тем, чтобы внешние раны оставались чистыми, чтобы свести к минимуму инфекцию. Это можно сделать с помощью спирта, перекиси водорода или просто воды с мылом.
Остальная часть этого раздела посвящена различным бактериям, которые, как известно, вызывают ЯФ.
Стрептококки группы А (Streptococcus pyogenes)
Бактерии стрептококка группы А являются одними из наиболее частых причин НФ. Эти грамположительные бактерии обычно переносятся людьми через нос, горло и кожу без каких-либо симптомов. Существует более 100 типов (серотипов) стрептококка группы А. Стрептококковая инфекция — это бактерия, вызывающая стрептококковое воспаление горла. Однако при попадании в организм он может вызвать некротический фасциит. Когда стрептококк группы А вызывает НФ, это может происходить вместе со стрептококковым токсическим шоковым синдромом (STSS), который увеличивает скорость и смертельность инфекции.
Золотистый стафилококк
Бактерии золотистого стафилококка становятся все более частой причиной НФ. Как и Strep, эти грамположительные бактерии обычно переносятся людьми через нашу кожу или нос без каких-либо симптомов. Бактерии стафилококка могут вызывать фурункулы и пищевые отравления. Метициллин-устойчивый золотистый стафилококк (MRSA) — это штамм этих бактерий, который является основным источником внутрибольничных инфекций, но за последнее десятилетие стал все более распространенным среди населения.Он также часто встречается в общественных местах, таких как раздевалки, общежития и дома престарелых. Его устойчивость к антибиотикам вызывает беспокойство во время лечения, но для лечения MRSA доступны специальные антибиотики.
Клебсиелла
Клебсиелла — грамотрицательные бактерии, обычно встречающиеся в природе. Известно, что они вызывают пневмонию, инфекции мочевыводящих путей, а также НФ.
Escherichia coli
Escherichia coli — грамотрицательные бактерии, часто встречающиеся в толстом кишечнике.Большинство из них безвредны, но, как известно, вызывают пищевое отравление. Большинство штаммов полезны для людей, когда они ограничены толстой кишкой (толстой кишкой), поскольку они обеспечивают витамин К и предотвращают распространение вредных бактерий в кишечнике. Однако E. Coli вне кишечника может вызвать инфекцию, в том числе NF.
Bacteroides
Bacteroides — анаэробные (могут выжить без кислорода) грамотрицательные бактерии, которые обычно находятся во рту, кишечнике и гениталиях. Эти бактерии обычно приносят пользу людям, предотвращая колонизацию кишечника серьезными патогенами.Однако, когда бактероиды инфицируют фасцию (например, через перфорированный кишечник), может возникнуть НФ.
Clostridium
Clostridium — анаэробные грамположительные бактерии, которые обычно встречаются в почве, а также в кишечнике людей и животных. Clostridium botulinum — это бактерия, вызывающая ботулизм, но существует множество других типов Clostridium, включая Clostridium perfringens,
Clostridium histolylicum, Clostridium septicum и Clostridium sordellii. Эти бактерии не так часто вызывают ЯБ, но, как известно, вызывают ЯБ и газовую гангрену.
Pseudomonas
Pseudomonas — грамотрицательные бактерии, широко распространенные в природе.