Что скрывается под буквами ЭДТА?
Есть ли на свете более загадочная аббревиатура, чем EDTA (далее -ЭДТА)? Наверняка, но сегодня мы с помощью инстаграм-блогера Ксении Кузнецовой расскажем именно об этих четырёх буквах, которые так часто встречаются в составах привычных косметических средств.
ЭДТА – это этилендиаминтетрауксусная кислота. Она имеет кристаллическую форму и добывается синтетическим путём. Основное её свойство – хелатирующее. Да-да, имено с “Х”! Эта кислота прекрасно связывает ионы металлов. Кстати, именно за это качество ЭДТА так любят в медицине. Её назначают пациентам (в 5% растворе) в рамках хелатирующей терапии, когда необходимо связать все вредные вещества в организме. Попадая в кровь, раствор ЭДТА обволакивает ионы тяжёлых металлов (даже таких, как ртуть и свинец), препятствуя их взаимодействию с клетками, способствуя их естественному “бегству” из организма. Вещество используется для нормализации уровня кальция и магния. Учёные также отмечают положительное влияние ЭДТА при атеросклерозе и нарушенном мозговом кровообращении у пожилых людей. А ещё эту кислоту применяют в стоматологии для размягчения некоторых тканей. Кроме того, она усиливает их проницаемость. Это необходимо при обработке узких корневых каналов в зубе.
А ещё эту кислоту применяют в стоматологии для размягчения некоторых тканей. Кроме того, она усиливает их проницаемость. Это необходимо при обработке узких корневых каналов в зубе.
Однако применяют ЭДТА и её производные далеко не только в медицине. Любят её и в пищевой промышленности, в качестве добавки Е-385. Загляните в свой холодильник и поизучайте этикетки майонеза, кетчупа, да и других продуктов. Есть немалая вероятность того, что в составе найдётся эта добавка. “Зачем же нас без нашего ведома пичкают лекарством?” Про хелатирующее свойство ещё не забыли? Кислота связывает металлы в продуктах, тем самым препятствуя их окислению, а, значит, продлевая срок годности.
Помимо пищевой промышленности, это вещество часто добавляют и в рецептуры косметических средств, ведь оно усиливает действие остальных ингредиентов. Например, способствует образованию густой пены.
Как распознать на этикетке?
Аналитическая химия насчитывает больше 60 веществ, в формуле которых есть ЭДТА. Но мы не будем углубляться в химию и просто поможем вам понять, на что стоит обращать внимание, изучая состав. Во-первых, 4 буквы – ЭДТА (EDTA). Это могут быть Sodium/Disodium/Trisodium/
Но мы не будем углубляться в химию и просто поможем вам понять, на что стоит обращать внимание, изучая состав. Во-первых, 4 буквы – ЭДТА (EDTA). Это могут быть Sodium/Disodium/Trisodium/
Опасность ЭДТА и веществ, её содержащих
Несмотря на многие громкие заявления, достоверных сведений о канцерогенности этой группы веществ я не обнаружила. Такие источники как CIR (Cosmetic Ingredient Review) и EWG’s Skin Deep характеризуют уровень опасности как низкий. Один из крупнейших порталов научных публикаций в области медицины PubMed предоставляет исследование о безопасности использования ЭДТА в косметических средствах со следующими оговорками:
1) Минимальная доза вещества, вызывающая токсикологическую реакцию у животных – 750мг/кг/день.
2) Клинические испытания не выявили поглощения солей ЭДТА через кожу.
Однако было установлено, что они истончают мембрану клетки, тем самым улучшая проницаемость, а, значит, потенциальное проникновение иных вредных веществ в организм. Кроме того, ЭДТА оказывает токсичное воздействие на организм при регулярном контакте с аэрозолями (освежители воздуха, средства для укладки, духи и т.п.)
3) Несмотря на то, что 95% ЭДТА и производных выводятся из организма в течение суток, оставшиеся 5% имеют свойство накапливаться в организме, а большое количество этих соединений оказывает цитотоксическое действие (подавляет работу клеток).
Пена для ванны “Солнышко” состав
Ещё один повод задуматься о целесообразности использования средств, содержащих ЭДТА и её производные, – их воздействие на окружающую среду. Как они туда попадают? Очень просто – шампуни, гели для душа, мытья посуды и пр. По данным Федерации Европейских микробиологических обществ (FEMS), соли ЭДТА растворяются в грунтовых водах, что в долгосрочной перспективе может повлиять на водные организмы, посредством изменения pH водоёмов. В зависимости от разновидности, соли ЭДТА могут разлагаться в природе от 15 часов до нескольких десятков и сотен лет.
В зависимости от разновидности, соли ЭДТА могут разлагаться в природе от 15 часов до нескольких десятков и сотен лет.
Наконец, международные экостандарты для натуральной косметики, в том числе, COSMOS Standard, категорически запрещают использование ЭДТА. Однако в рамках российской системы добровольной экологической сертификации бытовой химиии «Листок жизни» это вещество и его производные допускаются. Это неприятное для себя открытие я сделала, вчитавшись в микроскопическими буквами написанный состав средства для мытья посуды BioMio, сертифицированный “Листком жизни”.
Есть ли альтернатива?
Использование ЭДТА в косметических средствах и бытовой химии уже длительное время ставится под вопрос экспертами европейской косметической промышленности. Так, натуральной альтернативой ЭДТА является фитиновая кислота, получаемая (внимание!) из рисовой шелухи. А для образования приятной пены существует масса мягких растительных тензидов.
Ну что, друзья, время проверять свои баночки и бутылёчки?
Автор: Ксения Кузнецова, инстаблогер
ЭДТА — это.
 .. Что такое ЭДТА?
.. Что такое ЭДТА?ЭДТА — ЭДТУ металл ЭДТУ хелаты Этилендиаминтетрауксусная кислота (ЭДТУ), комплексон II: белый мелкокристаллический порошок, малорастворим в воде, нерастворим в большинстве органических растворителей, растворим в щелочах. ЭДТУ четырехосновная кислота,… … Википедия
ЭДТА — этилендиаминтетраацетат … Словарь сокращений русского языка
ЭДТА — сокр. от этилендиаминтетрауксусная кислота … Химические термины
Трис-ацетатный-ЭДТА буфер TAE — Трис ацетатный ЭДТА буфер, TAE * трыс ацэтатны ЭДТА буфер, ТАЕ * trisacetate EDTA buffer or TAE буфер, широко используемый для электрофоретического разделения молекул () ДНК или РНК в агарозном или полиакриламидном геле. Его состав: 40 ммоль Tris … Генетика. Энциклопедический словарь
Растворимость натриевых солей ЭДТА в зависимости от температуры (г/100 мл воды) — Степень замещения протонов ЭДТА на ионы натрия 22 °С 40 °С 80 °С Nah4edta 1,4 1,4 … Химический справочник
Константы диссоциации ЭДТА (lg K) — Протонированная форма комплексона 25 °С и μ=0,1 20 °С и μ=0,1 20 °С и μ=0 [Hedta]/[edta][H] 10,17 10,24 11,014 … Химический справочник
Константы устойчивости lg K комплексов ЭДТА с металлами главной подгруппы второй группы Периодической системы (при 20 °С и μ=0,1) — Катион K=[ML]/[M][L] K=[HML]/[H][ML] K=[HML]/[M][HL] Be2+ 9,2 Mg2+ 8 … Химический справочник
Коэффициенты разделения пар редкоземельных элементов, полученных при использовании ЭДТА в качестве элюента — РЗЭ α Lu Yb 1,9 Yb Tu 1,8 Er Ho 1,8 … Химический справочник
Логарифмы констант образования комплексов с ЭДТА при 20° и ионной силе 0,1 (KNO3 или KCl)
Логарифмы коэффициентов побочных реакций αY = [Y’]/[Y4-] для аниона Y4- этилендиаминтетрауксусной кислоты (ЭДТА) — pH lg αY 0 21,4 0,5 19,41 1,0 17,44 … Химический справочник
ЭДТА в составе микроудобрений – новая угроза для урожайности и плодородия почв.
 ,
,Среди веществ, наличие которых должно насторожить, часто встречается ЭДТА, предупреждает журналист, эколог Анна Козельская
На правах спонсорства
Покупая продукты в магазинах, мы все чаще обращаем внимание на их состав, учимся расшифровывать аббревиатуры, напечатанные мелкими буквами на упаковках. Потому что знаем: среди консервантов, красителей и прочих веществ, закодированных таким образом, могут содержаться и вредные для здоровья. Но не менее внимательно нужно относиться и к аббревиатурам, указанным на упаковках удобрений. Ведь тут действует то же правило, что и в отношении пищевых продуктов: не навреди.
С наступлением очередного сезона полевых работ, множество агрономов, фермеров и просто дачников пополняют свои запасы удобрений и средств защиты растений. В последние годы они все чаще покупают хелатные микроудобрения, применение которых – удобный, эффективный и экономически оправданный способ обеспечить растения необходимыми элементами питания.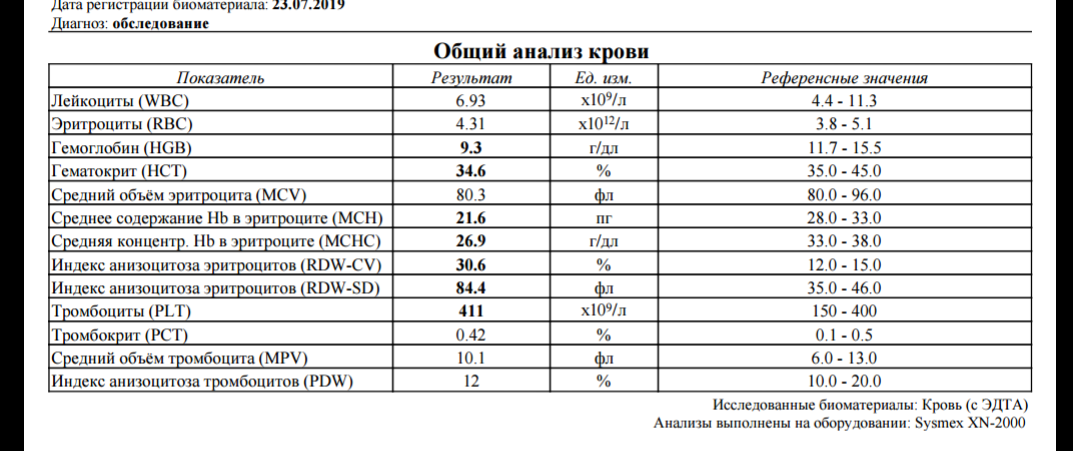 Их преимущества уже оценили миллионы аграриев. Предложений таких препаратов на украинском рынке – множество. К сожалению, среди них немало и таких, которые могут не только принести пользу, но и нанести в перспективе серьезный ущерб. Среди веществ, наличие которых должно насторожить, часто встречается ЭДТА.
Их преимущества уже оценили миллионы аграриев. Предложений таких препаратов на украинском рынке – множество. К сожалению, среди них немало и таких, которые могут не только принести пользу, но и нанести в перспективе серьезный ущерб. Среди веществ, наличие которых должно насторожить, часто встречается ЭДТА.
Прежде всего – безопасность
Какие бы препараты не планировал покупать сельхозпроизводитель – удобрения или средства защиты растений – перед ним постоянно стоит нелегкий выбор, ведь всегда есть желание приобрести именно тот продукт, который будет эффективным, безвредным, да еще и недорогим. И тут важно правильно расставить приоритеты. Конечно, все химические вещества, которые человек использует в сельском хозяйстве, пищевой промышленности и т.д., должны быть нетоксичными, не накапливаться в почве и живых организмах. Отклонение от этих требований обязательно приведет через некоторое время к серьезным проблемам. И, сэкономив один раз небольшую сумму на покупке дешевого удобрения или пестицида, в будущем можно получить гораздо более серьезные убытки.
Транспортное средство для микроэлементов
Требования, касающиеся безопасности для человека и окружающей среды, в полной мере относятся и к используемым в производстве микроудобрений веществам – комплексонам. Чтобы растение могло легко поглотить и усвоить те или иные микроэлементы, они должны быть внесены в биологически активной форме. Иными словами, необходимо своеобразное «транспортное средство», которое доставит нужный элемент в растение и «отдаст» его. Таким удобным «транспортом» оказались биологически активные комплексоны, или хелатирующие агенты.
Производители микроудобрений используют в своих производственных циклах различные хелатирующие агенты, одним из которых является ЭДТА (EDTA).
Именно на ее основе сделано подавляющее большинство микроудобрений, которые продают в Украине. Распространены они и в других странах. Поэтому ученые изучают этот комплексон наиболее тщательно. Итак, что скрыто за такой непонятной рядовому украинскому фермеру аббревиатурой как ЭДТА?
ЭДТА – это органическое соединение (этилендиаминтетрауксусная кислота), которое впервые было получено австрийским химиком Фердинандом Мюнцем в Германии еще в 1935 году.
Основное свойство ЭДТА – это способность связывать ионы некоторых элементов, среди которых Са 2+, Fe 3+, Cu 2+, Ni 2+, Со 2+, Zn 2+. Она образует с ними особенно стабильные комплексы, которые назвали хелатами (от греческого chele — крабовая клешня). Это свойство, а также относительно невысокая стоимость хелатанта, обусловили его широкое использование в различных сферах деятельности человека, таких как аналитическая химия, медицина, промышленность, сельское хозяйство. Поэтому ежегодный объем мирового производства ЭДТА составляет более 100 тыс. тонн.
Влияние ЭДТА на почвы и культурные растения
Конечно, аграриев прежде всего интересует влияние удобрений с ЭДТА на растения, почвы и эффективность их применения. И вот здесь как раз и кроется множество проблем, которые проявляются не сразу.
В частности, доказано, что микроудобрения на основе ЭДТА непригодны для корневых подкормок на карбонатных и щелочных почвах (pH> 8).
ЭДТА способна отнимать кальций из любых структур, его содержащих, в том числе и из растительных клеток. А кальций – важный мезоэлемент, который участвует в ключевых физиологических и биохимических процессах, протекающих в живом организме. Внешне дефицит элемента может проявляться как увядание растений. Недостаток кальция приводит к снижению урожайности, ухудшению качества семян и плодов, уменьшению сроков их хранения. Поэтому, после внесения микроудобрений на основе ЭДТА, желательно проводить внекорневую обработку растений удобрениями, содержащими кальций.
Кроме того, ЭДТА не усваивается растением. Отдав растению микроэлемент, хелатант попадает в почву, концентрируясь в основном в ее верхнем десятисантиметровом слое. Так как ЭДТА очень устойчивое соединение, то, оказавшись в земле, не разрушается и может связывать тяжелые металлы (в том числе ртуть, мышьяк, кадмий, свинец и другие). Тяжелые металлы, находящиеся в почве в виде солей, обычно плохо усваиваются растениями. Образование же комплексов ЭДТА с тяжелыми металлами приводит к более интенсивному и неконтролируемому их поглощению корневой системой.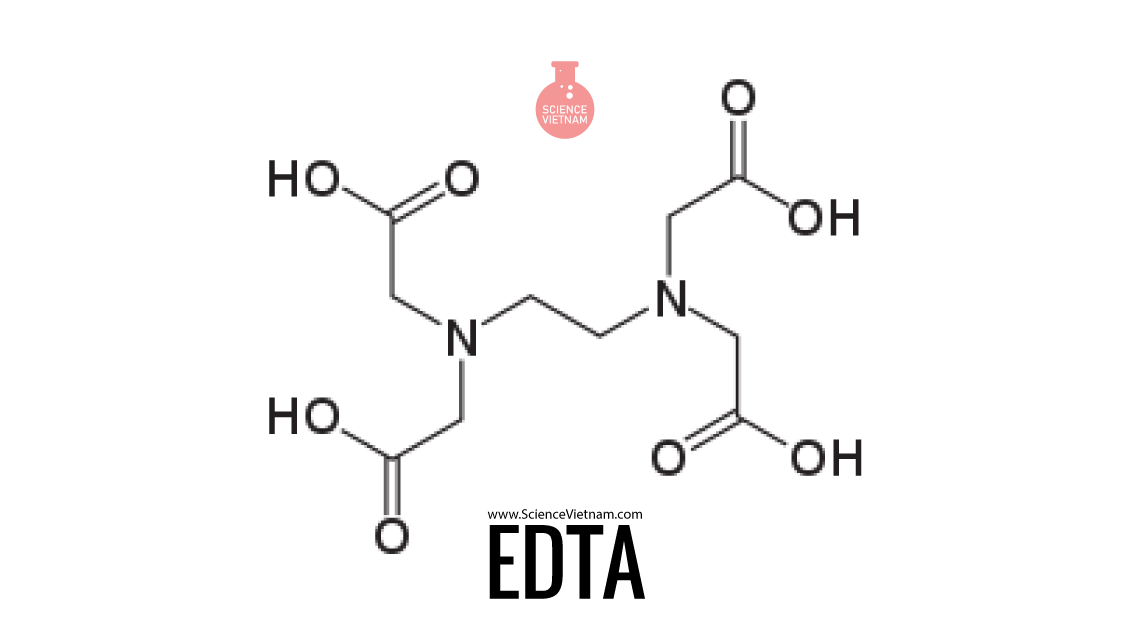 Исследования показывают, что ЭДТА влияет на накопление растениями металлов даже на третий год после ее однократного внесения. В результате концентрация тяжелых металлов в растениях может повышаться до токсичного уровня, что приводит к их угнетению, уменьшению урожайности и снижению качества продукции.
Исследования показывают, что ЭДТА влияет на накопление растениями металлов даже на третий год после ее однократного внесения. В результате концентрация тяжелых металлов в растениях может повышаться до токсичного уровня, что приводит к их угнетению, уменьшению урожайности и снижению качества продукции.
ЭДТА и окружающая среда
Масштабное применение ЭДТА повышает риск ее негативного воздействия на окружающую среду, так как это химическое соединение плохо разрушается в природе. Поэтому за достаточно короткое время оно стало одним из самых распространенных антропогенных загрязнителей.
Включаясь в естественный круговорот веществ, ЭДТА постепенно попадает в природные водоемы, а со временем накапливается в мировом океане, где переводит отложения токсичных металлов в растворимые соединения. Это вызывает отравления планктона, рыб, птиц и высших животных. Примером экологической катастрофы, произошедшей в результате неконтролируемого применения ЭДТА, может служить ситуация, сложившаяся в дельте реки Миссисипи в США.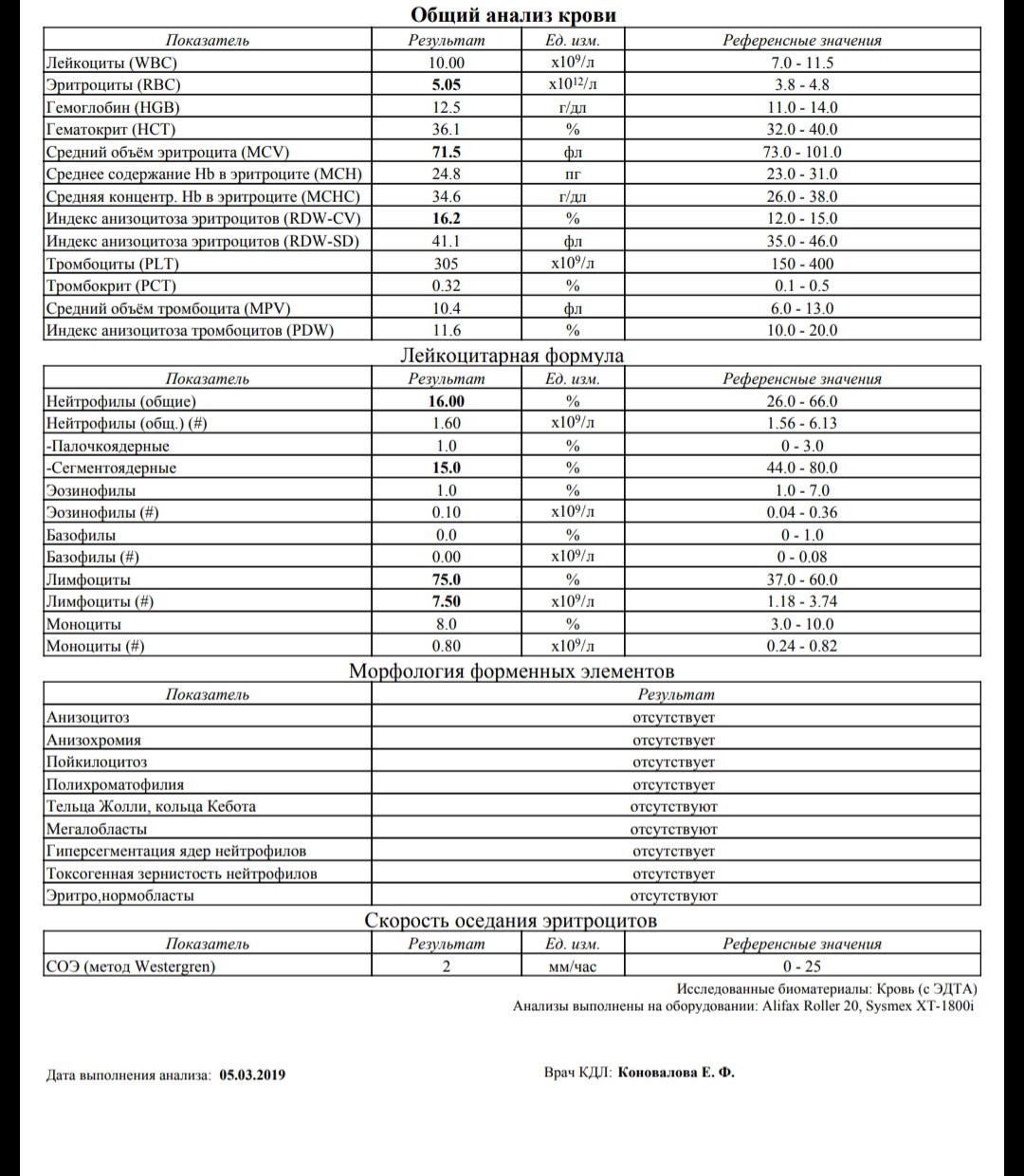 Там сейчас течет поток, «обогащенный» ЭДТА и другими химическими веществами, способствующими увеличению ежегодной «мертвой зоны», которая охватывает огромную территорию.
Там сейчас течет поток, «обогащенный» ЭДТА и другими химическими веществами, способствующими увеличению ежегодной «мертвой зоны», которая охватывает огромную территорию.
Таким образом, использование соединений на основе ЭДТА является потенциально опасным для сельского хозяйства и окружающей среды. В долгосрочной перспективе оно приводит к увеличению загрязнения сельскохозяйственных земель, деградации почв, уменьшению урожайности и ухудшению качества сельскохозяйственной продукции.
Исправить эти последствия очень трудно, а иногда – и невозможно. Стоит вспомнить лишь о популярном когда-то препарате ДДТ, больше известном как просто «дуст». Из-за выраженной токсичности и способности сохраняться в окружающей среде, ДДТ запретили во всем мире и не используют уже полвека. Но до сих пор его находят в почвах и живых организмах в различных частях нашей планеты, даже в ледниках Антарктиды.
Для тех, кто хочет больше узнать о свойствах и использовании ЭДТА и других комплексонов, рекомендуем ознакомиться с книгами и статьями ученых, исследующих это вещество: В. Васильева, С. Чугаева, А. Бучинской, А. Кульчинской, Л. Мартыненко, Н. Кузьминой, Ю. Самусенко, И. Автуховича, Д. Постникова, О. Четверик, Е. Груздевой, В. Архиповой и других. Все эти материалы можно найти в свободном доступе в Интернете.
Васильева, С. Чугаева, А. Бучинской, А. Кульчинской, Л. Мартыненко, Н. Кузьминой, Ю. Самусенко, И. Автуховича, Д. Постникова, О. Четверик, Е. Груздевой, В. Архиповой и других. Все эти материалы можно найти в свободном доступе в Интернете.
Автор: Журналист, эколог Анна Козельская
Лейкоцитарная формула (Differential White Blood Cell Count, лейкограмма)
Процентное соотношение различных видов лейкоцитов (нейтрофилов, лимфоцитов, моноцитов, эозинофилов, базофилов) периферической крови.
Кроме того, лейкоциты различаются по степени зрелости. Большая часть клеток-предшественников зрелых форм лейкоцитов (юные, миелоциты, промиелоциты, бластные формы клеток), а также плазматические клетки, молодые ядерные клетки эритроидного ряда и др. в периферической крови появляются только в случае патологии.
При многих тяжелых инфекциях, септических и гнойных процессах лейкоцитарная формула меняется за счет увеличения количества палочкоядерных нейтрофилов, а также возможного появления более юных форм — метамиелоцитов и миелоцитов. Такое изменение лейкограммы с увеличением процентного содержания молодых форм нейтрофилов называют сдвигом влево; увеличение же в основном за счет сегментоядерных и полисегментоядерных форм (возникающее при мегалобластной анемии, болезнях почек и печени, состоянии после переливания крови) – называют сдвигом вправо. Значительное омоложение клеток (в крови отмечается присутствие метамиелоцитов, миелоцитов, промиелоцитов, бластных клеток) может указывать на хронические лейкозы, эритролейкоз, миелофиброз, метастазы злокачественных новообразований, острые лейкозы. В таких случаях принято говорить о лейкемоидной реакции.
Такое изменение лейкограммы с увеличением процентного содержания молодых форм нейтрофилов называют сдвигом влево; увеличение же в основном за счет сегментоядерных и полисегментоядерных форм (возникающее при мегалобластной анемии, болезнях почек и печени, состоянии после переливания крови) – называют сдвигом вправо. Значительное омоложение клеток (в крови отмечается присутствие метамиелоцитов, миелоцитов, промиелоцитов, бластных клеток) может указывать на хронические лейкозы, эритролейкоз, миелофиброз, метастазы злокачественных новообразований, острые лейкозы. В таких случаях принято говорить о лейкемоидной реакции.
Лейкоцитарная формула отражает относительное (процентное) содержание лейкоцитов различных видов, и увеличение или снижение процентного содержания лимфоцитов может носить как абсолютный, так и относительный характер.
В лейкоцитарную формулу входят следующие параметры:
- Лимфоциты
- Сегментоядерные нейтрофилы
- Палочкоядерные нейтрофил
- Бластные клетки
- Промиелоциты
- Миелоциты
- Метамиелоциты
- Моноциты
- Базофилы
- Эозинофилы
- Плазматические клетки
Лимфоциты (LYM — lymphocytes)
Гетерогенная популяция клеток, образующихся в костном мозге.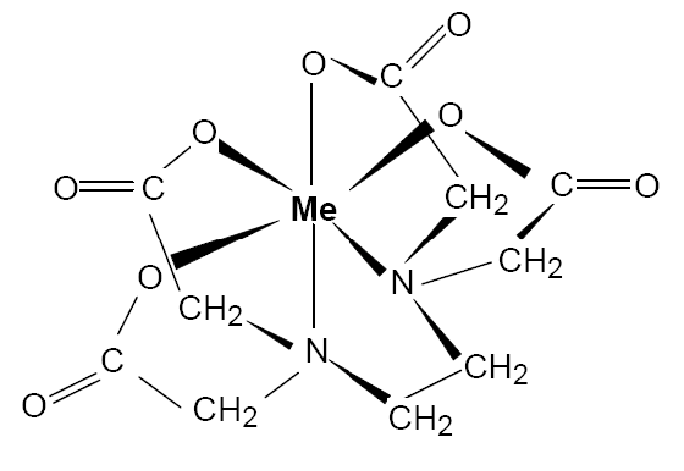 Основная функция — реализация иммунного ответа. Т-лимфоциты определяют клеточный иммунитет, выполняют регуляторные и эффекторные функции. В-лимфоциты участвуют в гуморальном иммунитете. Референсные значения (вариант нормы) Лимфоциты (LYM — lymphocytes) -абсолютное содержание, 109 клеток /л
Основная функция — реализация иммунного ответа. Т-лимфоциты определяют клеточный иммунитет, выполняют регуляторные и эффекторные функции. В-лимфоциты участвуют в гуморальном иммунитете. Референсные значения (вариант нормы) Лимфоциты (LYM — lymphocytes) -абсолютное содержание, 109 клеток /л| Возраст | Мужчины | Женщины |
|---|---|---|
| 3 мес – 1 год | 3,3 – 9,0 | |
| 1 год — 6 лет | 1,6 – 7,1 | |
| 6 лет — 11 лет | 0,9 – 5,0 | |
| 11 лет — 17 лет | 1,1 – 3,8 | |
| > 17 лет | 1,26 – 3,35 | |
| Возраст | Мужчины | Женщины |
|---|---|---|
| <2 нед | 22-55 | |
| 2 нед – 1 год | 45-70 | |
| 1–2 года | 37-60 | |
| 2 года – 5 лет | 33-55 | |
| 5–9 лет | 30-50 | |
| 9–15 лет | 30-46 | |
| >15 лет | 19-37 | |
| Увеличение значений (лимфоцитоз) | Уменьшение значений (лимфоцитопения) |
|---|---|
Реактивный:
|
|
Моноциты (MONO — monocytes)
Самые крупные клетки периферической крови.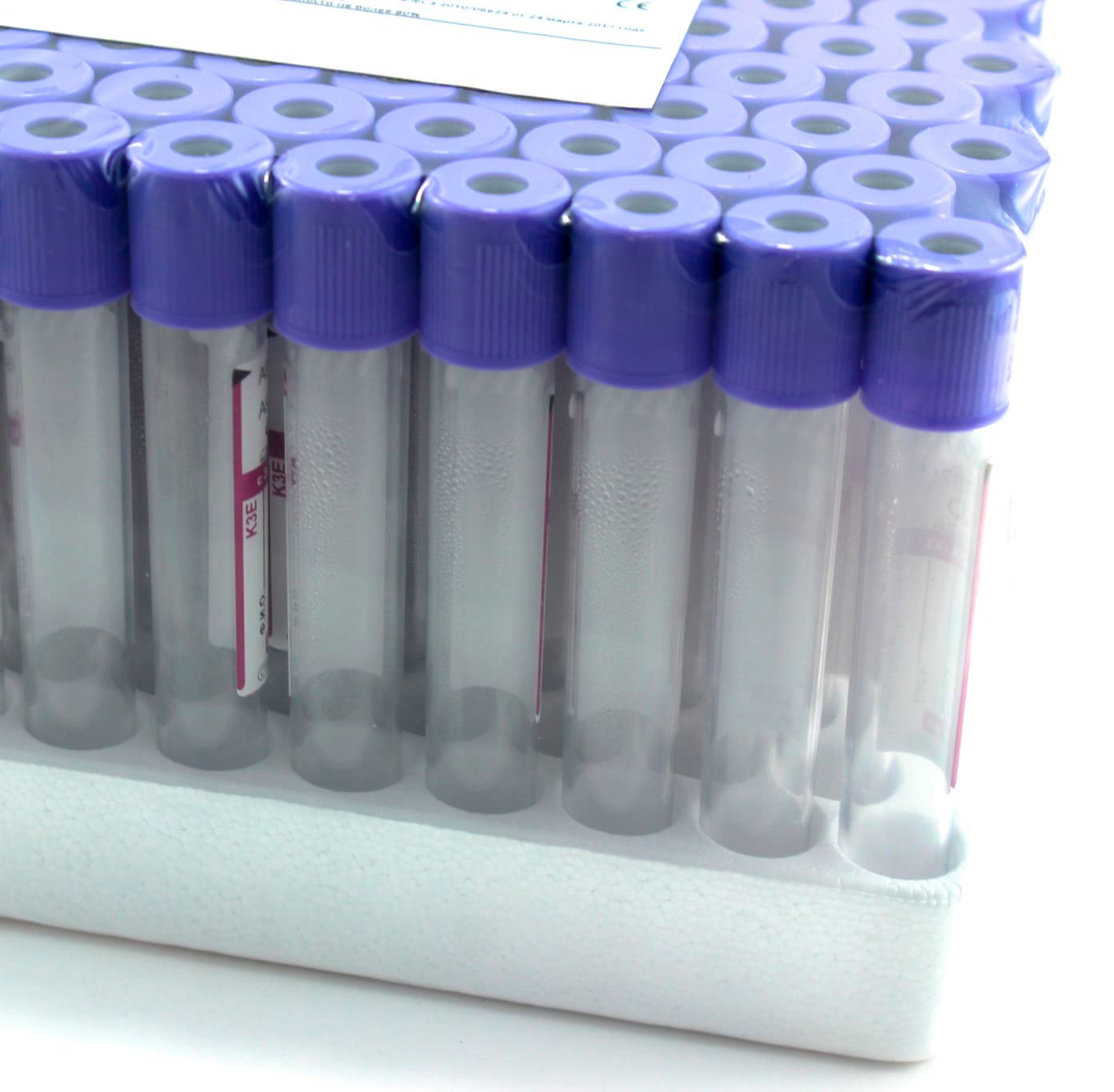 Моноциты обладают выраженной фагоцитарной функцией (являются макрофагами), участвуют в защитных реакциях организма путем продукции цитокинов, в процессах обмена веществ.
Моноциты обладают выраженной фагоцитарной функцией (являются макрофагами), участвуют в защитных реакциях организма путем продукции цитокинов, в процессах обмена веществ.
Референсные значения (вариант нормы)
Моноциты (MONO — monocytes) — %
| Возраст | Мужчины | Женщины |
|---|---|---|
| <2 нед | 5,0–15,0 | |
| 2 нед – 1 год | 4,0–10,0 | |
| 1–2 года | 3,0–10,0 | |
| 2 года – 15 лет | 3,0–9,0 | |
| >15 лет | 3,0–11,0 | |
Моноциты (MONO — monocytes) -абсолютное содержание, 109 клеток /л
| Возраст | Мужчины | Женщины |
|---|---|---|
| 3 мес – 17 лет | 0,37 – 1,26 | |
| > 17 лет | 0,29 – 0,95 | 0,25 – 0,84 |
| Увеличение значений (моноцитоз) | Уменьшение значений (моноцитопения) |
|---|---|
Реактивный:
Опухолевой:
|
|
Пробирка вакуумная с ЭДТА К2 и К3
Пробирки IMPROVACUTER содержат в качестве антикоагулянта EDTA-K2 или EDTE-K3 (этилендиаминтетраацетат)
Вакуумные пробирки Improvacuter для гематологический исследований с К2 и К3 ЭДТА. Применяются при необходимости лабораторных исследований цельной крови.
Применяются при необходимости лабораторных исследований цельной крови.
Область применения: гематология, ПЦР- диагностика, исследование групп крови, большее применение находят в клинической гематологии. Пробирки с EDTA подходят для различных анализаторов клеток крови. ЭДТА связывает ионы кальция и блокирует каскад реакций свертывания крови. При этом концентрация и характеристики клеточных и внеклеточных компонентов остаются практически неизменными.
Основные характеристики:
Кодовый цвет крышки: фиолетовый
Материал пробирки: пластик
Материал для исследования: цельная кровь
Применение: гематология
Наполнители: EDTA K2, EDTA K3
Размер пробирок (мм.): 13×75, 13×100
Стандартный объем образца (мл): 1, 2, 3, 4, 5, 6
Сухая или жидкая ЭДТА K3 или ЭДТА K2 нанесена на внутреннюю стенку пробирки в концентрации 1.2 мг — 2.0 мг (0.00411 моль/литр — 0.006843 моль/литр) сухой ЭДТА на 1 мл крови.
Гематологические пробирки подходят для использования в автоматических анализаторах без открывания крышки. Игла анализатора легко протыкает резиновую часть пробки пробирки.
Игла анализатора легко протыкает резиновую часть пробки пробирки.
Рекомендуемая скорость центрифугирования: 3500 об/мин.
Максимальная скорость центрифугирования: 5000 об/мин.
Рабочая температура: 0ºС-37ºС
Максимальное отклонение содержания вакуума: 10%
Упаковка:
13 мм — 100 штук в штативе / 1000 штук (10 штативов) в коробке
16 мм — 50 штук в штативе / 1000 штук (20 штативов) в коробке
Для гематологических исследований (ЭДТА-К3)
EDTA K3 Tube
| Артикул | Цвет крышки | Объем взятия крови, мл | Размер, мм | Цена за 1 шт. | Тип пробки | Упаковка |
| 692021112 | фиолетовый | 2 мл | 13х75 мм | 6,00 | SCA | 100/1000 |
| 692031112 | 3 мл | 13х75 мм | SCA | 100/1000 | ||
| 692041112 | 4 мл | 13х75 мм | SCA | 100/1000 | ||
| 692051112 | 5 мл | 13х75 мм | SCA | 100/1000 | ||
| 693031112* | 3 мл | 13х100 мм | SCA | 100/1000 | ||
| 693041112* | 4 мл | 13х100 мм | SCA | 100/1000 | ||
| 693051112* | 5 мл | 13х100 мм | SCA | 100/1000 | ||
| 693061112 | 6 мл | 13х100 мм | SCA | 100/1001 | ||
| 694081212* | 8 мл | 16х100 мм | 8,00 | SCB | 50/1000 | |
| 694091212 | 9 мл | 16х100 мм | SCB | 50/1000 |
* — отмечены товары по пред заказу — срок ожидания от 3 до 5 месяцев
Для гематологических исследований (ЭДТА-К2)
EDTA K2 Tube
| Артикул | Цвет крышки | Объем взятия крови, мл | Размер, мм | Цена за 1 шт. | Тип пробки | Упаковка |
| 682011112 | фиолетовый | 1 мл | 13х75 мм | 6,00 | SCA | 100/1000 |
| 682021112 | 2 мл | 13х75 мм | SCA | 100/1000 | ||
| 682031112 | 3 мл | 13х75 мм | SCA | 100/1000 | ||
| 682041112 | 4 мл | 13х75 мм | SCA | 100/1000 | ||
| 683031112 | 3 мл | 13х100 мм | SCA | 100/1000 | ||
| 683041112* | 4 мл | 13х100 мм | SCA | 100/1000 | ||
| 683051112* | 5 мл | 13х100 мм | SCA | 100/1000 | ||
| 684081212* | 8 мл | 16х100 мм | 8,00 | SCB | 50/1000 |
* — отмечены товары по пред заказу — срок ожидания от 3 до 5 месяцев
С данным товаром также покупают:
Обратная транскрипция
Прайминг обратной транскрипции
Выбор праймеров, которые будут использоваться для инициации обратной транскрипции, может сильно повлиять на результаты RT-qPCR.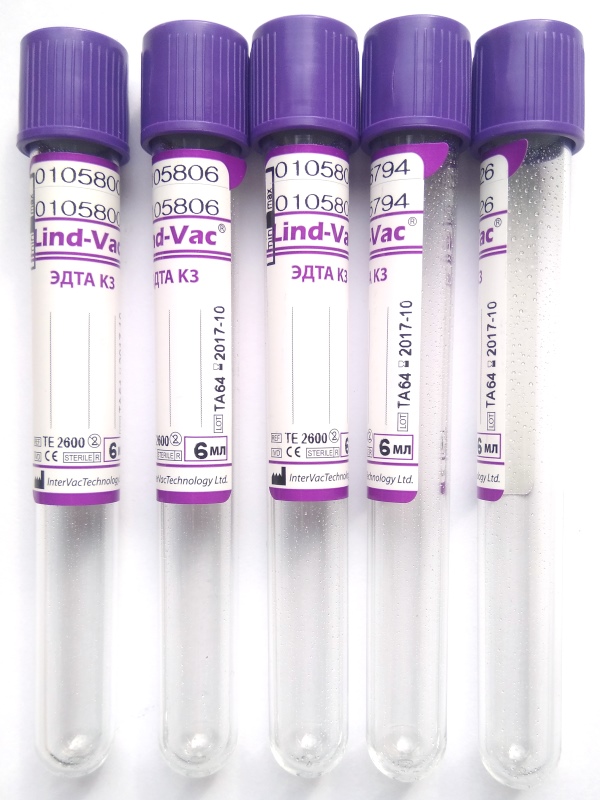 Для одноступенчатого
Для одноступенчатого
RT-qPCR, используются специфичные для генов праймеры. При выполнении двухэтапного анализа можно использовать обратный ген-специфический праймер, олиго-dT (O4387), случайные гексамеры, нонамеры, декамеры, додекамеры или пентадекамеры 2 или комбинацию олиго-dT (O4387) и случайных праймеров. использовал. Ген-специфическое праймирование обычно выполняется в отдельных реакциях для каждой целевой РНК.Эти отдельные реакции могут иметь очень разную эффективность, что затрудняет сравнение концентраций РНК. С другой стороны, при использовании специфичных для генов праймеров весь продукт RT будет кодировать интересующий ген и может позволить количественное определение мРНК с очень низким содержанием, которые не могут быть обнаружены с помощью неспецифических праймеров RT. Чтобы избежать потенциально высоких вариаций RT между анализами, которые могут возникать с ген-специфическими праймерами, можно использовать неспецифические праймеры для создания пула кДНК. Отдельные анализы qPCR для каждой мишени могут быть выполнены впоследствии с аликвотами из пула кДНК. Если все мишени кПЦР находятся рядом с 3’-концом полиаденилированной мРНК, подходящим выбором праймера является олиго-dT (O4387). С другой стороны, если цели кПЦР находятся на расстоянии более нескольких килобаз от 3’-конца или если РНК не полиаденилирована, случайные праймеры приведут к более надежному обнаружению. Если относительное 3’-расположение мишеней кПЦР варьируется или желаемые транскрипты содержат комбинацию полиаденилированных и неполиаденилированных РНК, смесь олиго-dT (O4387) и случайных олигомеров даст наилучшие результаты
Отдельные анализы qPCR для каждой мишени могут быть выполнены впоследствии с аликвотами из пула кДНК. Если все мишени кПЦР находятся рядом с 3’-концом полиаденилированной мРНК, подходящим выбором праймера является олиго-dT (O4387). С другой стороны, если цели кПЦР находятся на расстоянии более нескольких килобаз от 3’-конца или если РНК не полиаденилирована, случайные праймеры приведут к более надежному обнаружению. Если относительное 3’-расположение мишеней кПЦР варьируется или желаемые транскрипты содержат комбинацию полиаденилированных и неполиаденилированных РНК, смесь олиго-dT (O4387) и случайных олигомеров даст наилучшие результаты
Праймирование обратной транскрипции для двухэтапных реакций RT
Для двухэтапного RT-прайминга обычно используются два подхода; oligo-dT (O4387) и случайное праймирование.Метод олиго-dT (O4387) основан на гибридизации олиго-dT (обычно 15-мерного) с поли-A-хвостом, присутствующим на 3 ’конце большинства молекул мРНК, для праймирования и выборочной обратной транскрипции мРНК.
Этот подход, хотя концептуально очень простой, имеет связанные проблемы: связывание праймера олиго-dT (O4387) неспецифично для мРНК при температурах реакции, используемых для RT, таким образом, олиго-dT (O4387) будет связываться неспецифически с другими областями. РНК. Кроме того, участки рРНК также обнаруживаются, поскольку области ATrich в этих молекулах примированы олиго-dT (O4387).Некоторые мРНК, например мРНК, кодирующие гистоны, не содержат поли-A-хвостов и могут не быть представлены в полученной кДНК.
Второй метод использует случайное заполнение. Случайные праймеры состоят из случайных последовательностей, часто из гексамеров (6-мер) или нонамеров (9-мер). Они используются для инициации реакции RT, что приводит к синтезу фрагментов кДНК различной длины, которые представляют собой исходную РНК. Случайные праймеры гибридизуются по длине транскрипта и имеют тенденцию быть более толерантными к вторичной структуре, чем олиго-dT (O4387) или ген-специфическое праймирование.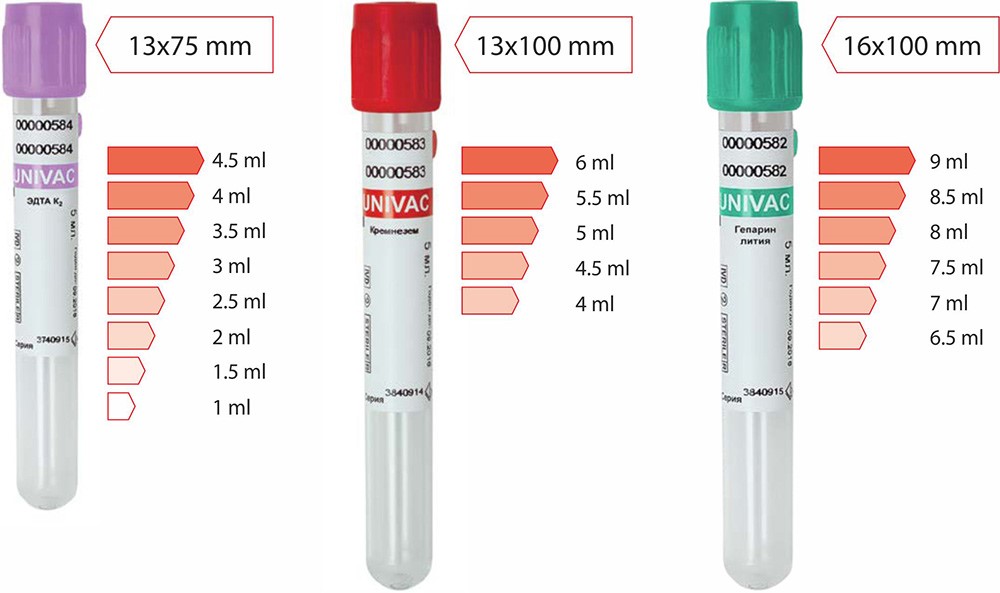
Чтобы воспользоваться преимуществами соответствующих методов, некоторые протоколы требуют комбинации обоих типов праймеров. Конкретный праймер для целевой последовательности также может использоваться в двухэтапном протоколе RT, но чаще используется в одноэтапной процедуре (см. Ниже).
Праймирование обратной транскрипции для одноэтапных реакций RT
В одноэтапном протоколе RT используются специфичные для генов праймеры для обратной транскрипции единственной мишени. Дизайн геноспецифического праймера имеет решающее значение; он должен находиться в открытой доступной области мРНК-мишени, если прогнозируется при температуре реакции RT.В этих условиях существует линейная зависимость между входной РНК и кДНК (рис. 8.4). Этот праймер может быть (и обычно является) общим для праймера ПЦР.
Экспрессия РНК-транскрипта фактора вирулентности в подроде Leishmania Viannia: влияние вида, источника изолята и вируса РНК-1 Leishmania
Fraga J, Veland N, Montalvo AM, Praet N, Boggild AK, Valencia BM, Arevalo J , Llanos-Cuentas A, Dujardin JC, Van der Auwera G. Точное и быстрое типирование видов из кожных и кожно-слизистых лейшманиозных поражений в Новом Свете.Диагностика Microbiol Infect Dis. 2012; 74: 142–50.
Точное и быстрое типирование видов из кожных и кожно-слизистых лейшманиозных поражений в Новом Свете.Диагностика Microbiol Infect Dis. 2012; 74: 142–50.
CAS Статья Google Scholar
Рейтинджер Р., Дюжарден Дж. С., Лузир Х, Пирмез К., Александр Б., Брукер С. Кожный лейшманиоз. Lancet Infect Dis. 2007. 7 (9): 581–96.
Артикул Google Scholar
Olivier M, Atayde VD, Isnard A, Hassani K, Shio MT. Leishmania Факторы вирулентности: основное внимание уделяется металлопротеиназе GP63.Микробы заражают. 2012. 14 (15): 1377–89.
CAS Статья Google Scholar
Бифельд Э., Клос Дж. Генетика вирулентности Leishmania . Med Microbiol Immunol. 2015; 204: 619–34.
CAS Статья Google Scholar
Ламотт С.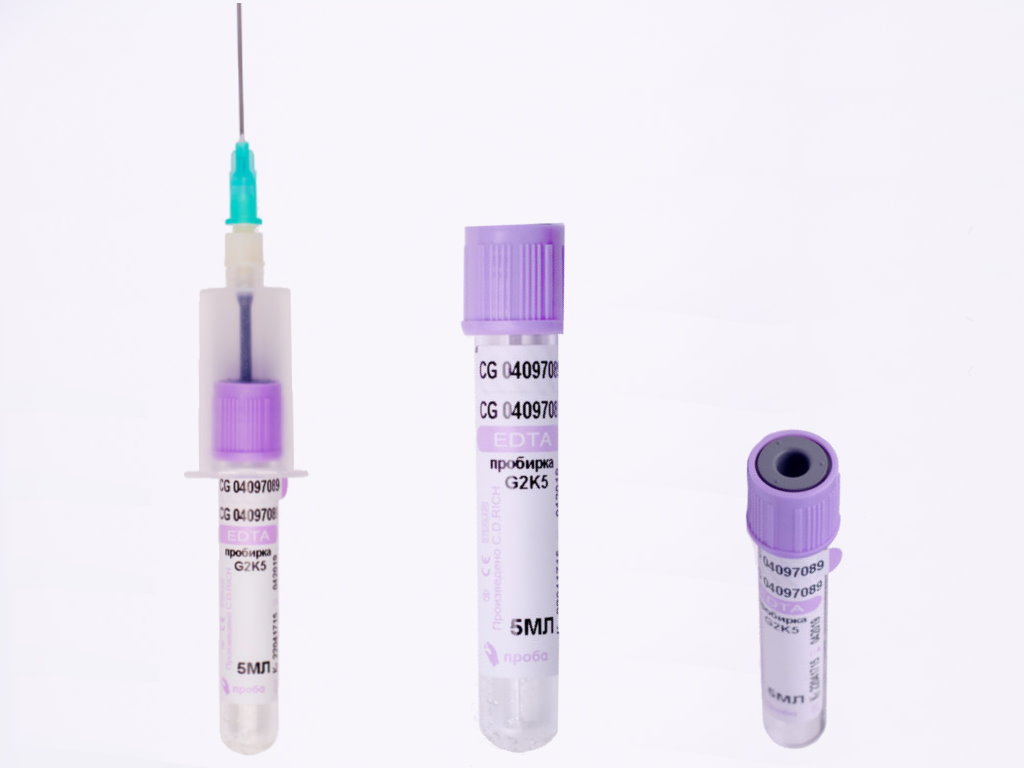 , Спат Г.Ф., Прина Э. Враг внутри: нацеливание на взаимодействие паразита и хозяина для открытия антилейшманиозных препаратов.PLoS Negl Trop Dis. 2017; 11 (6): e0005480.
, Спат Г.Ф., Прина Э. Враг внутри: нацеливание на взаимодействие паразита и хозяина для открытия антилейшманиозных препаратов.PLoS Negl Trop Dis. 2017; 11 (6): e0005480.
Артикул Google Scholar
Аронсон Н., Хервальдт Б.Л., Либман М., Пирсон Р., Лопес-Велес Р., Вейна П., Карвалью Е.М., Эфрос М., Джеронимо С., Мэджилл А. Диагностика и лечение лейшманиоза: руководство по клинической практике, подготовленное Институтом инфекционных заболеваний. Американское общество болезней (IDSA) и Американское общество тропической медицины и гигиены (ASTMH). Clin Infect Dis. 2016; 63 (12): e202–64.
Артикул Google Scholar
Hartley MA, Ronet C, Zangger H, Beverley SM, Fasel N. Leishmania РНК-вирус: когда хозяин оплачивает пошлину. Cell Infect Microbiol. 2012; 2: 99.
Google Scholar
Ginouves M, Simon S, Bourreau E, Lacoste V, Ronet C, Couppie P, Nacher M, Demar M, Prevot G. Распространенность и распространение Leishmania RNA вируса 1 в Leishmania паразитов из Франции Гвиана. Am J Trop Med Hyg. 2016; 94 (1): 102–6.
Распространенность и распространение Leishmania RNA вируса 1 в Leishmania паразитов из Франции Гвиана. Am J Trop Med Hyg. 2016; 94 (1): 102–6.
CAS Статья Google Scholar
Маседо Д.Х., Менезес-Нето А., Ругани Д.М., Роча А.С., Сильва С.О., Мело М.Н., Лай Л.Ф., Беверли С.М., Гонтихо С., Соарес Р.П. Низкая частота LRV1 у штаммов Leishmania braziliensis , выделенных из типичных и атипичных поражений в штате Минас-Жерайс, Бразилия. Мол Биохим Паразитол. 2016; 210 (1–2): 50–4.
CAS Статья Google Scholar
Pereira L, Maretti-Mira AC, Rodrigues KM, Lima RB, Oliveira-Neto M, Cupolillo E, Pirmez C, Oliveira MP. Тяжесть тегументарного лейшманиоза не связана исключительно с инфекцией вируса 1 РНК Leishmania в Бразилии. Mem Inst Oswaldo Cruz. 2013; 108 (5): 665–7.
CAS Статья Google Scholar
Ives A, Ronet C, Prevel F, Ruzzante G, Fuertes-Marraco S, Schutz F, Zangger H, Revaz-Breton M, Lye LF, Hickerson SM, Beverley SM, Acha-Orbea H, Launois P , Фазель Н., Масина С. Leishmania РНК-вирус контролирует тяжесть кожно-слизистого лейшманиоза. Наука. 2011; 331 (6018): 775–8.
CAS Статья Google Scholar
Ронет С., Беверли С.М., Фазель Н. Слизисто-кожный лейшманиоз в Новом Свете. Landes Biosci. 2011; 2 (6): 547–52.
Google Scholar
Валенсия Б.М., Адауи V, Чантри М., Альба М., Рамос А., Аревало Дж., Льянос-Куэтас А., Боггильд А.К.Спорная роль вируса РНК Leishmania как детерминанта патогенности лейшманиоза человека. Int J Infect Dis. 2014; 21С: 165–6.
Артикул Google Scholar
Ogg MM, Carrion R, de Carvalho Botelho AC, Mayrink W, Correa-Oliveira R, Patterson JL. Количественная оценка РНК вируса Leishmania в клинических образцах и ее возможная роль в патогенезе. Am J Trop Med Hyg. 2003. 69: 309–13.
Количественная оценка РНК вируса Leishmania в клинических образцах и ее возможная роль в патогенезе. Am J Trop Med Hyg. 2003. 69: 309–13.
Артикул Google Scholar
Bourreau E, Ginouves M, Prevot G, Hartley MA, Gangneux P, Robert-Gangneux F, Dufour J, Marie DS, Bertolotti A, Pratlong F, Martin R, Schutz F, Couppie P, Fasel N, Ronet C. Leishmania — Наличие вируса РНК в L . guyanensis увеличивает риск неэффективности лечения первой линии и симптоматического рецидива. J Infect Dis. 2016; 213: 105–11.
CAS Статья Google Scholar
Atayde VD, Hassani K, Filho ASL, Borges AR, Adhikari A, Martel C, Olivier M. Leishmania экзосомы и другие факторы вирулентности: влияние на врожденный иммунный ответ и функции макрофагов. Cell Immunol. 2016; 309: 7–18.
CAS Статья Google Scholar
Сулат Д., Бодган С. Функция макрофагальных и паразитарных фосфатаз при лейшманиозе. Фронт Иммунол. 2017; 8: 1838.
Артикул Google Scholar
Sutter A, Antunes D, Silva-Almeida M, Costa MGS, Caffarena ER.Структурное понимание лейшманолизинов, кодируемых на хромосоме 10 на Leishmania ( Viannia ) braziliensis . Mem Inst Oswaldo Cruz. 2017; 112 (9): 617–25.
CAS Статья Google Scholar
МакКонвилл М.Дж., де Соуза Д., Сондерс Э., Ликич В.А., Надерер Т. Жизнь в фаголизосоме; метаболизм амастигот Leishmania . Trends Parasitol. 2007. 23 (8): 368–71.
CAS Статья Google Scholar
Дункан Р.К., Салотра П., Гоял Н., Акопянц Н.С., Беверли С.М., Нахаси Х.Л. Применение технологии микрочипов экспрессии генов для исследования кинетопластид. Curr Mol Med. 2004; 4: 611–21.
2004; 4: 611–21.
CAS Статья Google Scholar
Holzer TR, McMaster WR, Forney JD. Профилирование экспрессии с помощью полногеномной межвидовой гибридизации микроматрицы выявляет дифференциальную экспрессию генов в проциклических промастиготах, амастиготах, полученных из повреждений, и аксенических амастиготах в Leishmania mexicana .Мол Биохим Паразитол. 2006; 146: 198–218.
CAS Статья Google Scholar
Leifso K, Cohen-Freue G, Dogra N, Murray A, McMaster WR. Геномный и протеомный анализ экспрессии промастигот Leishmania и стадий жизни амастигот: геном Leishmania экспрессируется конститутивно. Мол Биохим Паразитол. 2007. 152: 35–46.
CAS Статья Google Scholar
Cohen-Freue G, Holzer TR, Forney JD, McMaster WR. Глобальная экспрессия генов в Leishmania . Int J Parasitol. 2007; 37: 1077–86.
Int J Parasitol. 2007; 37: 1077–86.
CAS Статья Google Scholar
McNicoll F, Drummelsmith J, Muller M, Madore E, Boilard N, et al. Комбинированный протеомный и транскриптомный подход к изучению дифференцировки стадий у Leishmania infantum . Протеомика. 2006; 6: 3567–81.
CAS Статья Google Scholar
Depledge DP, Evans KJ, Ivens AC, Aziz N, Maroof A, et al. Сравнительное профилирование экспрессии Leishmania : модуляция экспрессии генов между видами и в различных генетических фонах хозяина. PLoS Negl Trop Dis. 2009; 3 (7): e476.
Артикул Google Scholar
Rastrojo A, Carrasco-Ramiro F, Martin D, Crespillo A, Reguera RM, Aguado B, Requena JM. Транскриптом Leishmania major на стадии аксенической промастиготы: аннотация транскрипта и относительные уровни экспрессии с помощью RNA-seq. BMC Genomics. 2013; 14: 223.
BMC Genomics. 2013; 14: 223.
CAS Статья Google Scholar
Кристенсен С.М., Диллон ЛАЛ, Карвалью Л.П., Пассос С., Новаис Ф.О., Хьюитт В.К. и др. Метатранскриптомное профилирование кожного поражения человека — Leishmania braziliensis . PLoS Negl Trop Dis. 2016; 10 (9): e0004992.
Артикул Google Scholar
Кариявасам Р., Гревал Дж., Лау Р., Пурсел А., Валенсия Б.М., Льянос-Куэтас А., Боггилд А.К.Влияние Leishmania РНК вируса 1 на экспрессию провоспалительных биомаркеров в модели макрофагов человека американского тегументарного лейшманиоза. J Infect Dis. 2017; 216 (7): 877–66.
CAS Статья Google Scholar
Zangger H, Ronet C, Desponds C, Kuhlmann FM, Robinson J, Hartley MA, Prevel F, Castiglioni P, Pratlong F, Bastien P, Müller N, Parmentier L, Saravia NG, Beverley SM, Fasel N Обнаружение РНК вируса Leishmania у паразитов Leishmania .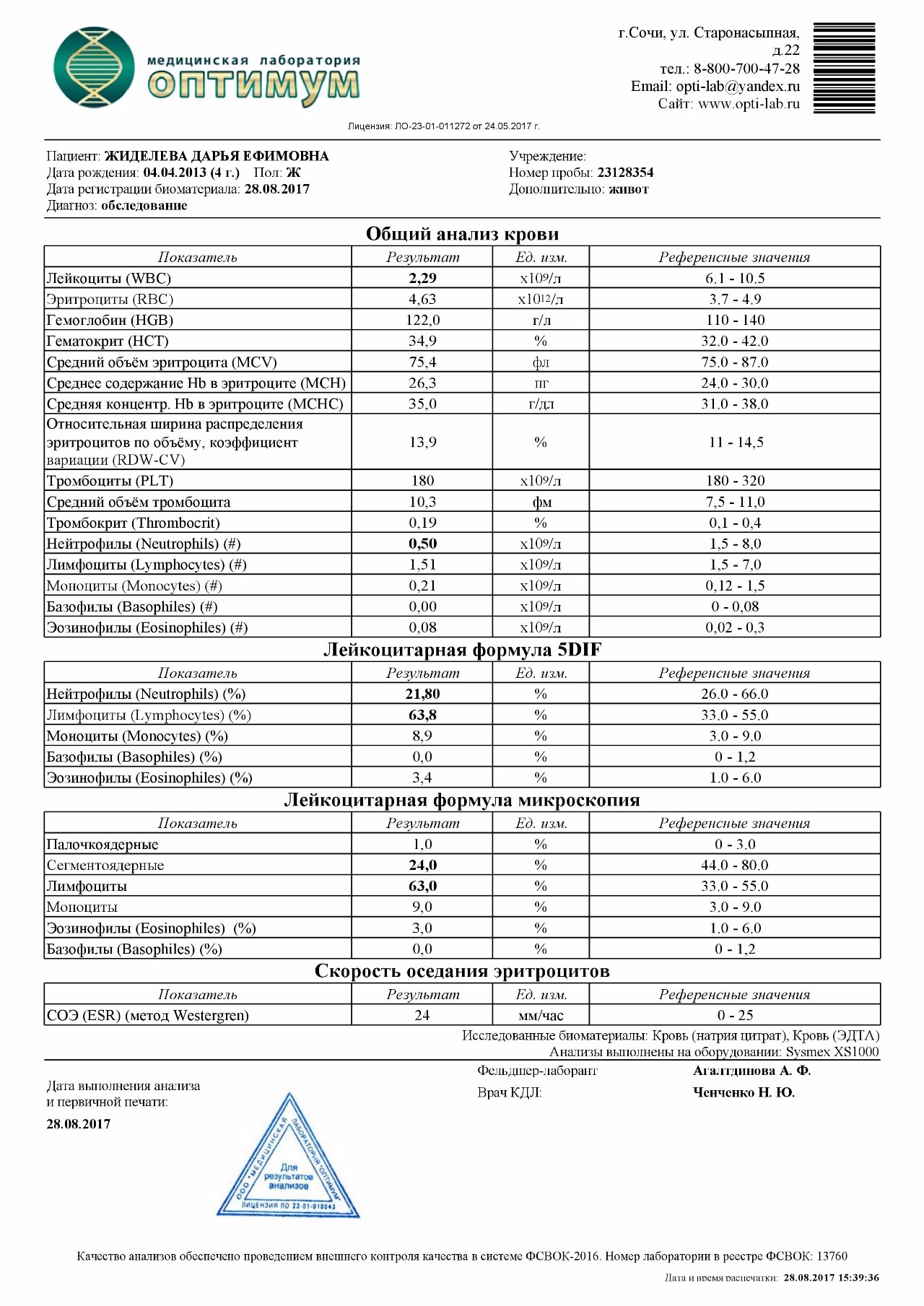 PLoS Negl Trop Dis. 2013; 7 (1): e2006.
PLoS Negl Trop Dis. 2013; 7 (1): e2006.
CAS Статья Google Scholar
Schmittgen TD, Livak KJ. Анализ данных ПЦР в реальном времени методом сравнительной КТ. Nat Protoc. 2008; 3: 1101–8.
CAS Статья Google Scholar
Рекена Дж. М., Монтальво А. М., Фрага Дж. Молекулярные шапероны Leishmania: центральные игроки во многих связанных со стрессом и не связанных физиологических процессах.Biomed Res Int. 2015; 2015: 301326.
Артикул Google Scholar
Hombach A, Ommen G, MacDonald A, Clos J. Небольшой белок теплового шока необходим для термотолерантности и внутриклеточного выживания Leishmaina donovani . J Cell Sci. 2014; 127 (Pt 21): 4762–73.
Артикул Google Scholar
Drini S, Criscuolo A, Lechat P, Imamura H, Skalicky T. , Rachidi N, Lukes J, Dujardin JC, Spath GF.Видовая и штаммоспецифическая адаптация суперсемейства HSP70 у патогенных трипаносоматид. Genome Biol Evol. 2016; 8 (6): 1980–95.
, Rachidi N, Lukes J, Dujardin JC, Spath GF.Видовая и штаммоспецифическая адаптация суперсемейства HSP70 у патогенных трипаносоматид. Genome Biol Evol. 2016; 8 (6): 1980–95.
CAS Статья Google Scholar
Кробич С., Брандау С., Хойер С., Шметц С., Хьюбел А., Клос Дж. Leishmania donovani белок теплового шока 100. Характеристика и функция при дифференциации состояния амастигот. J Biol Chem. 1998. 273 (11): 6488–94.
CAS Статья Google Scholar
Holakuyee M, Mahdavi M, Hassan ZM, Abolhassani M. Обогащенные белками теплового шока — промастиготы Leishmania major , индуцирующие иммунный ответ Th3 у мышей BALB / c. Иран Биомед Дж. 2012; 16 (4): 209–17.
CAS PubMed PubMed Central Google Scholar
Parra-Munoz M, Aponte S, Ovalle-Bracho C, Saavendra CH, Echeverry MC. К объективной диагностике для обнаружения вируса РНК Leishmania (LRV) в клинических образцах от пациентов с кожным лейшманиозом.BioRxiv: 338095. https://doi.org/10.1101/338095.
К объективной диагностике для обнаружения вируса РНК Leishmania (LRV) в клинических образцах от пациентов с кожным лейшманиозом.BioRxiv: 338095. https://doi.org/10.1101/338095.
Mottram JC, Coombs GH, Alexander J. Цистеиновые пептидазы как факторы вирулентности Leishmania . Curr Opin Microbiol. 2004. 7 (4): 357–81.
Артикул Google Scholar
Casgrain PA, Martel C, McMaster WR, Mottram JC, Olivier M, Descoteaux A. Цистеинпептидаза B регулирует вирулентность Leishmania mexicana посредством модуляции экспрессии GP63.PLoS Pathog. 2016; 12 (5): e1005658.
Артикул Google Scholar
Pereira BAS, Britto C, Alves CR. Экспрессия генов, связанных с инфекцией, у паразитов и хозяина во время экспериментального заражения мышей Leishmania ( Leishmania ) amazonensis . Microb Pathog. 2012. 52 (2): 101–8.
CAS Статья Google Scholar
Silva-Almeida M, Pereira BAS, Ribeiro-Guimaraes ML, Alves CR. Протеиназы как факторы вирулентности у Leishmania spp. инфекция у млекопитающих. Векторы паразитов. 2012; 5: 160.
CAS Статья Google Scholar
Гарами А., Илг Т. Роль изомеразы фосфоманнозы в синтезе и вирулентности гликоконъюгатов Leishmania mexicana . J Biol Chem. 2001. 276 (9): 6566–75.
CAS Статья Google Scholar
Brandau S, Dresel A, Clos J. Высокие конститутивные уровни белков теплового шока у патогенных паразитов человека из рода Leishmania . Biochem J. 1995; 310: 225–23247.
CAS Статья Google Scholar
Fernandes MC, Dillon LAL, Belew AT, Bravo HC, Mosser DM, El-Sayed NM. Двойное транскриптомное профилирование человеческих макрофагов, инфицированных Leishmania , выявляет различные сигнатуры репрограммирования. mBio.2016; 7 (3): e00027–16.
Двойное транскриптомное профилирование человеческих макрофагов, инфицированных Leishmania , выявляет различные сигнатуры репрограммирования. mBio.2016; 7 (3): e00027–16.
CAS Статья Google Scholar
| Тип: | Дефицитные продукты (плазма и сыворотка) |
| Номер каталога: | D250005 |
| Разновидность: | Человек |
| Формат: | Плазма |
| Размер: | 10 х 1 мл |
| Область исследования: | Диабет, нефротический синдром, болезнь сердца, метаболизм, рак, гипоксия |
| Энтрез Джин: | 9370 |
| Омим: | 605441 |
| UniProt: | Q15848 |
| Unigene: | Hs. 80485 80485 |
| Синонимы: | Связанный с комплементом адипоцит белок 30 кДа Связанный с комплементом адипоцит белок 30 кДа ACDC ACRP 30 ACRP30 ADIPO_HUMAN Адипоцит Адипоцит C1q и домен коллагена, содержащий белок Адипоцит, связанный с комплементом Белок 30 кДа Адипоцит, связанный с комплементом белок 30 кДа Адипоцитный белок, связанный с комплементом 30 кДа Адипоцитный белок, связанный с комплементом 30 кДа, специфичный для адипоцитов, связанный с комплементом белок Адипонектин Предшественник адипонектина адипонектин, C1Q и домен коллагена, содержащий AdipoQ Транскрипт гена, наиболее распространенного в жировой ткани 1, транскрипт гена, наиболее распространенного в жировой ткани 1, белок, специфичный для жировой ткани, коллаген, подобный фактору ADIPQTL1, ADPN, APM 1, apM-1, ApM1, C1q и белок, содержащий домен коллагена, GBP 2828 Связывание с желатином белок Желатин-связывающий белок 28 Желатин-связывающий белок |
Хелатирующие лиганды: определение, модели и примеры — стенограмма видео и урока
Номенклатура — правила и примеры
Хелатирующие комплексы могут быть, ну, сложными, поэтому иногда необходимы специальные префиксы, чтобы избежать путаницы. Префиксы бис-, трис-, тетракис- и т.д. заменяются на ди-, три-, тетра- и т.д. Названия лигандов затем помещаются в скобки после префикса.
Префиксы бис-, трис-, тетракис- и т.д. заменяются на ди-, три-, тетра- и т.д. Названия лигандов затем помещаются в скобки после префикса.
Хорошим примером является диметиламин. Если бы мы не использовали бис- и трис-, диметиламин мог быть либо «двумя метилами, связанными с амином ((Ch4) 2 NH)», либо «двумя метиламинами ((Ch4 Nh3) 2)». Очевидно, этого не будет, поэтому мы используем диметиламин для обозначения «два метила, один амин» и бис (метиламин) для обозначения «двух метиламинов».»
Будучи полезными в любом потенциально неоднозначном комплексе, бис- и трис- всегда используются, когда ди-, три- и т. Д. Уже присутствуют в названии лиганда. Напоминаем, что лиганды названы в алфавитном порядке, а степень окисления металла указана в скобках.
Рассмотрим, например, [Co ((Nh3 Ch3 Ch3 Nh3) 2 Cl2] + — дихлорбис (этилендиамин) кобальт (III). В этом комплексе присутствуют два лиганда: анионный лиганд хлора и нейтральный хелатирующий лиганд этилендиамин.Само название «этилендиамин» содержит термин «ди-», поэтому «два этилендиаминовых лиганда» записывается как «бис (этилендиамин)».
Хелатирующие лиганды, которые имеют анионные донорные участки, могут одновременно удовлетворять вторичной валентности (координационный центр) и первичной валентности (заряд) на ионе металла. Такие хелатные комплексы называются внутренними металлическими комплексами или внутренними комплексами.
Комплексы называются внутренними комплексами первого порядка , если координационное число и заряд иона металла точно уравновешиваются лигандом.Они нейтральны. Давайте посмотрим на бис (ацетилацетонато) дигидроксоNi (II), чтобы понять, что это означает. Ацетилацетон (acac) представляет собой бидентатный анионный лиганд, который образует шестичленное хелатное кольцо с ионом металла. Как и другие переходные металлы, Ni имеет координационное число (или вторичную валентность) 6; здесь он имеет заряд +2 (это его первичная валентность), поэтому первичная валентность металла удовлетворяется двумя анионными лигандами acac, а вторичная валентность удовлетворяется шестью связями.
Лиганды с анионными донорными центрами могут также образовывать комплексы, в которых общий анионный заряд лиганда не точно уравновешивает заряд на ионе металла.Такие комплексы с остаточным зарядом называются внутренними металлическими комплексами второго порядка . Некоторыми примерами являются Na + [(acac) Co (II)] и K + [(acac) Pt (II) Cl2]. В этих комплексах координационное число (вторичная валентность) металла удовлетворяется, но заряд (первичная валентность) металла не удовлетворяется.
Внутренние комплексы летучие и в основном нерастворимы в воде, но растворимы в органических растворителях. У них низкая температура плавления и насыщенный цвет. Эти свойства делают их очень полезными в аналитической химии.
Краткое содержание урока
Лиганды с несколькими донорными сайтами, которые способны образовывать пяти- или шестичленные стабильные кольца с металлами, называются хелатирующими лигандами . Некоторые распространенные хелатирующие лиганды включают этилендиамин (en), ацетилацетон (acac) и этилендиаминтетрауксусную кислоту (EDTA). Когда первичная валентность (заряд) и вторичная валентность (координационное число) металла удовлетворены и комплекс является нейтральным, комплекс называется внутренним металлическим комплексом первого порядка .Когда первичная валентность (заряд) и вторичная валентность (координационное число) металла не удовлетворяются внутренне и комплекс имеет остаточный заряд, комплекс называется внутренним металлическим комплексом второго порядка .
Некоторые распространенные хелатирующие лиганды включают этилендиамин (en), ацетилацетон (acac) и этилендиаминтетрауксусную кислоту (EDTA). Когда первичная валентность (заряд) и вторичная валентность (координационное число) металла удовлетворены и комплекс является нейтральным, комплекс называется внутренним металлическим комплексом первого порядка .Когда первичная валентность (заряд) и вторичная валентность (координационное число) металла не удовлетворяются внутренне и комплекс имеет остаточный заряд, комплекс называется внутренним металлическим комплексом второго порядка .
Страница не найдена — PacBio
Соглашение об использовании изображения
Загружая, копируя или используя изображения, размещенные на этом веб-сайте («Сайт»), вы подтверждаете, что прочитали, поняли и согласны с условиями настоящего Соглашения об использовании изображений, а также с условиями, приведенными на веб-страницу Юридические уведомления, которые вместе регулируют использование вами изображений, как указано ниже.Если вы не согласны с такими условиями, не загружайте, не копируйте и не используйте изображения каким-либо образом, если у вас нет письменного разрешения, подписанного уполномоченным представителем Pacific Biosciences.
В соответствии с условиями настоящего Соглашения и условиями, приведенными на веб-странице Юридических уведомлений (в той степени, в которой они не противоречат условиям настоящего Соглашения), вы можете использовать изображения на Сайте исключительно для (а) редакционного использования в прессе. и / или отраслевых аналитиков, (б) в связи с обычной, рецензируемой, научной публикацией, книгой или презентацией и т.п.Вы не можете изменять или модифицировать любое изображение, полностью или частично, по любой причине. Вы не можете использовать какое-либо изображение таким образом, чтобы искажать представление о связанных продуктах, услугах или технологиях Pacific Biosciences или о любых связанных с ними характеристиках, данных или свойствах. Вы также не можете использовать какое-либо изображение таким образом, который обозначает какое-либо представление или гарантию (явную, подразумеваемую или установленную законом) от Pacific Biosciences в отношении продукта, услуги или технологии. Права, предоставляемые настоящим Соглашением, являются личными для вас и не могут быть переданы вами другой стороне.
Вы, а не Pacific Biosciences, несете ответственность за использование изображений. Вы признаете и соглашаетесь с тем, что любое неправильное использование изображений или нарушение настоящего Соглашения нанесет компании Pacific Biosciences непоправимый вред. Pacific Biosciences является владельцем или лицензиатом изображения, но не агентом владельца. Вы соглашаетесь предоставить Pacific Biosciences следующую кредитную линию: «Предоставлено Pacific Biosciences of California, Inc., Menlo Park, CA, USA», а также включаете любые другие кредиты или благодарности, отмеченные Pacific Biosciences.Вы должны включить любое уведомление об авторских правах, изначально включенное в изображения, на всех копиях.
ИЗОБРАЖЕНИЙ ПРЕДОСТАВЛЯЮТСЯ Pacific Biosciences «КАК ЕСТЬ». Pacific Biosciences ОТКАЗЫВАЕТСЯ ОТ ВСЕХ ЗАЯВЛЕНИЙ И ГАРАНТИЙ, ЯВНЫХ, ПОДРАЗУМЕВАЕМЫХ ИЛИ ЗАКОНОДАТЕЛЬНЫХ УСЛОВИЙ, ВКЛЮЧАЯ, НО НЕ ОГРАНИЧИВАЯСЬ НАРУШЕНИЕМ, СОБСТВЕННОСТЬЮ, КОММЕРЧЕСКОЙ ЦЕННОСТЬЮ И ПРИГОДНОСТЬЮ ДЛЯ ОПРЕДЕЛЕННОЙ ЦЕЛИ. НИ ПРИ КАКИХ ОБСТОЯТЕЛЬСТВАХ Pacific Biosciences НЕ НЕСЕТ ОТВЕТСТВЕННОСТИ ЗА ЛЮБЫЕ ПРЯМЫЕ, КОСВЕННЫЕ, СЛУЧАЙНЫЕ, КОСВЕННЫЕ ИЛИ КОСВЕННЫЕ УБЫТКИ ЛЮБОГО ТИПА, КАКИЕ-ЛИБО В ОТНОШЕНИИ ИЗОБРАЖЕНИЙ.
Вы соглашаетесь с тем, что Pacific Biosciences может прекратить ваш доступ и использование изображений, размещенных на веб-сайте PacificBiosciences.com, в любое время и без предварительного уведомления, если компания сочтет, что вы нарушили какое-либо из условий настоящего Соглашения об использовании изображений. Вы соглашаетесь возмещать, защищать и оградить Pacific Biosciences, ее должностных лиц, директоров, сотрудников, агентов, лицензиаров, поставщиков и любых сторонних поставщиков информации на Сайт от всех убытков, расходов, убытков и издержек, включая разумные гонорары адвокатам. в результате любого нарушения вами условий настоящего Соглашения об использовании изображений или прекращения Pacific Biosciences вашего доступа к Сайту или его использования.Прекращение действия не повлияет на права Pacific Biosciences или ваши обязательства, возникшие до прекращения.
РНК для трансляции в клетках для клинических исследований и иммунотерапии
Миссия CELLSCRIPT ™ заключается в предоставлении лучших продуктов и технологий для производства и использование РНК для трансляции в клетках для клинических исследований и терапии.
Текущие продукты включают наборы для транскрипции in vitro , Кэппирование 5 ‘РНК с использованием кэп-аналога или кэп-ферментов, и Полиаденилирование 3 ‘РНК, а также универсальные наборы для изготовления кеппированных, поли (A) -хвостовая мРНК для трансляции в клетках.
CELLSCRIPT ™ недавно представленные наборы для транскрипции РНК INCOGNITO ™ для in vitro синтез РНК, содержащей модифицированные нуклеозиды, такие как псевдоуридин (Ψ) и / или 5-метилцитидин (m5C) вместо соответствующие канонические нуклеозиды U или C (рис. 1). Эти комплекты названы так потому, что кэпированные, полиаденилированные и модифицированные нуклеозидами РНК-продукты (называемые «мРНК INCOGNITO») замаскированы таким образом, чтобы они не вызывают врожденных иммунные ответы в той же степени, что и соответствующие немодифицированные мРНК при трансфекции в клетки млекопитающих, которые экспрессируют различные Сенсоры РНК.
Как указано в журнальных статьях 1–3 и патентных заявках, 4 технологии, на которых используются наборы INCOGNITO RNA и другие продукты были изобретены профессорами Каталин Карико и Дрю Вайсман в Университет Пенсильвании и имеет эксклюзивную лицензию на CELLSCRIPT ™ для всех областей применения. Доктора Вайсман и Карико показали, что ИНКОГНИТО мРНК, помимо того, что она менее иммуногенна, транслируется в белок на гораздо более высокие уровни, чем соответствующая мРНК, не содержащая модифицированные нуклеозиды как в культивируемых клетках (рис. 2), так и в целом организмов (рисунок 3).Кодирование Ψ- и m5C-модифицированной мРНК типа INCOGNITO KLF4, LIN28, cMYC, OCT4 и SOX2, которые неоднократно трансфицировались в соматические клетки, такие как фибробласты и кератиноциты, приводили к высокому эффективное создание индуцированных плюрипотентных стволовых клеток. 5
Доставка кодирующей белок мРНК INCOGNITO в клетки в культуре или in vivo в организме может производить терапевтическое эффект за счет компенсации недостающего или дефектного белка, сверхэкспрессии желаемый белок, вызывающий изменение клеточного фенотипа или запускающий иммунный ответ, специфичный для заболевания.Таким образом, мы считаем, что мРНК INCOGNITO найдет множество применений в регенеративной медицине, например: клеточная перепрограммирование; клеточная терапия; трансплантация клеток, тканей или органов или ремонт; тканевая или органная инженерия; замена ферментов терапии; и иммунотерапия или терапия иммуномодуляции.
Связаться с CELLSCRIPT ™ для обсуждения наборов для проведения исследований или терапевтических оценок INCOGNITO мРНК или немодифицированной РНК, или возможное изготовление на заказ подходящего оценка и шкала РНК для вашего проекта.
Список литературы
- Karikó K, Buckstein M, Ni H, Weissman D. Иммунитет 23 : 165–75, 2005. Подавление распознавания РНК. Toll-подобных рецепторов: влияние модификации нуклеозидов и эволюционное происхождение РНК. Журнальная статья
- Карико К., Мурамацу Х, Валлийский Ф.А., Людвиг Дж., Като Х., Акира С., Вайсман Д. Молекулярная терапия 16 : 1833–40, 2008 г.Включение псевдоуридина в мРНК дает превосходные неиммуногенные вектор с повышенной трансляционной способностью и биологической стабильностью. Журнальная статья
- Андерсон Б. Мурамацу Х., Наллагатла С.Р., Бевилаква П. Сансинг L, Weissman D, Karikó K. Nucleic Acids Res 38 : 5884–92, 2010. Включение псевдоуридина в мРНК усиливает перевод за счет уменьшения активации PKR. Журнальная статья
- E.g., заявка на патент США № US 200
852 Журнальная статья
- Уоррен Л., Манос П.Д., Ахфельдт Т., Ло Й.Х., Ли Х., Лау Ф., Эбина В.,
Мандал П.К., Смит З.Д., Мейснер А., Дейли Г.К., Брак А.С., Коллинз Дж. Дж.,
Cowan C, Schlaeger TM, Rossi DJ. Cell Stem Cell. 7 (5) : 549–50, 2010.
Journal Статья .
EvidenceProf Blog
Этот пост является продолжением моего предыдущего поста (здесь) о тестировании ЭДТА в деле Стивена Эйвери / «Создание убийцы».Много было сказано о том факте, что «Марк ЛеБо, глава химического отдела в штаб-квартире ФБР в Вирджинии, … использовал … ЭДТА-тест, который ФБР использовало только однажды — в суде об убийстве [О.Дж.] Симпсона в 1995 году. . » Это буквально правда. Доказательства тестирования на ЭДТА до этого использовались только один раз. Но это была не единственная попытка представить такие доказательства.
В 2007 году другой житель Калифорнии попытался представить доказательства наличия ЭДТА. Это увлекательный случай, заслуживающий отдельного подкаста или серии Netflix.Я вернусь к его делу в следующих постах. Обращаясь к аргументу ответчика, суд отметил, что
На сегодняшний день единственным случаем, в котором были допущены результаты теста на ЭДТА, является Люди против Орентала Джеймса Симпсона …. Там д-р Баллард не принимал непосредственного участия в тестировании ЭДТА …. В единственном случае, когда Были предложены результаты тестирования доктора Балларда, Нью-Джерси против Джоша Помпея, Верховный суд Нью-Джерси обоснованно отверг как достоверность докт.Балларда и надежность тестирования ЭДТА в постановлении о неприемлемости свидетельства ЭДТА.
У меня нет доступа к фактам дела Помпея, но вот как суд описал
В единственном другом случае, когда рассматривалась допустимость результатов теста с ЭДТА, Нью-Джерси против Помпея, д-р Баллард провел тестирование и заявил, что обнаружил «значимые с судебной точки зрения» количества ЭДТА в нескольких пятнах крови. Однако суд Помпей категорически запретил докторуБалларда за то, что он не «предоставил [] простой научный контекст для измерения и сравнения своих результатов», оставив в суде «впечатление, что Баллард пропустил информацию, чтобы одновременно поддержать, преумножить и скрыть значение своих предположительно важных открытий ЭДТА . » [FN30] …. Суд Помпей обвинил доктора Балларда в «избирательном следовании научному методу» и в том, что он «имел план для достижения результатов, которые подтверждают теорию фальсификации ответчика.”…
Например, в Pompey, одно из пятен, в котором доктор Баллард обнаружил «повышенный» уровень ЭДТА, состояло исключительно из крови жертвы, которая никогда не хранилась в пробирке с пурпурным верхом … Балларда критиковали за совместное тестирование двух пятен, ни одно из которых не было связано с предполагаемым преступником, и за признание совокупного числа ЭДТА критически значимым. Баллард не нормализовал агрегированные числа для учета множественных пятен….
Назвав аналитические методы доктора Балларда «бессистемными и ненадежными», суд Помпей сделал следующие выводы:
В целом, он использовал достоверную науку (газовую хроматографию / масс-спектрометрию) для получения продукта, бойко и ненаучно отвергая ЭДТА из источников, отличных от пробирок с пурпурным верхом, и совершил гигантский шаг к выводу, который не поддерживается наукой, фактами в записи или даже здравом смысле.
Баллард исказил представление своих данных, скрыл значимость своих выводов и изменил свои гипотезы в соответствии с теорией фальсификации обвиняемого.Баллард не продемонстрировал, что его выводы основывались на надежных основаниях. Скорее, его постоянные двусмысленности дискредитировали его метод рассуждений и, таким образом, сделали его окончательные выводы бесполезными …
___________________
FN30. Суд Pompey был обеспокоен тем, что доктор Баллард не объяснил единицы измерения для своих различных расчетов. В этом случае д-р Баллард представил в учебном пособии определенные стандартные «концентрации» ЭДТА в крови. В отчете о его результатах заголовок «Концентрация ЭДТА» появляется без каких-либо дополнительных объяснений.Только заметив важное различие в единицах, представленных в учебном пособии (нг / л), и единицах, использованных в результатах доктора Балларда (нг / мл), Суд смог определить, что «концентрация ЭДТА», указанная в его результатах, обозначает концентрация ЭДТА в буферной жидкости, а не концентрация ЭДТА в крови.
Очевидно, что трудно сделать какие-либо твердые выводы, не зная всех фактов дела Помпея.