Что такое LM-активатор? статья пациентам «DENTAL PROGRESS»
LM-Activator™ является ортодонтическим аппаратом для раннего ортодонтического лечения, а также помогает зубам принять правильное положение при прорезывании. Стандартные аппараты имеют форму, соответствующую идеальной окклюзии, и выпускаются в разных размерах. 37 различных комбинаций моделей позволяют подобрать подходящий для конкретного пациента ЛМ Активатор без внесения индивидуальных изменений. Использование аппарата LM-Активатр способствует программированию правильного положения постоянных зубов при прорезывании, формированию физиологической окклюзии зубных рядов, оптимизирует функции мышц, рост челюстей, положение и артикуляцию языка.
ЛМ-Активаторы изготовлены из биосовместимого силикона. Жесткость материала подбиралась таким образом, чтобы трейнер не разжевывался и не вызывал сильного дискомфорта при ношении из-за чрезмерной жесткости аппарата
Маленькие пациенты быстро адаптируются к данному аппарату, так как LM-Активатор имеет увеличенную лингвальную кромку, что позволяет ему лучше удерживаться во рту во время сна; кроме того, трейнер имеет отверстия, которые дают возможность использовать аппарат даже при затрудненном носовом дыхании; силикон не изменяет цвет во время ношения.
Лечение c LM-активатором Время лечения. В большинстве случаев рекомендуется начинать лечение в раннем сменном прикусе до прорезывания верхних резцов. Наилучшие результаты при коррекции перекрестного прикуса достигаются в том случае, если лечение начинается на этапе прорезывания зубов. С учетом роста костей челюстно-лицевой области у детей в ходе лечения необходимо задействовать от 2 и более аппаратов.

Ретенционный период. Окончание этапа активного ношения зависит от индивидуальных особенностей пациента. В большинстве случаев лечение может быть завершено, когда нижние клыки занимают правильное положение и лингвальный ретейнер может быть закреплен на нижней дуге. Если у пациента есть предпосылки для ретрузии или глубокого прикуса, ночное ношение следует продолжать до окончания пубертатного периода роста. Во время ретенционного периода врача следует посещать каждые 6 месяцев. Любое ортодонтическое вмешательство должно быть основано на анализе зубочелюстной, скелетной и функциональной характеристик, а также с учетом индивидуальных характеристик роста пациента для обеспечения надлежащего и безопасного лечения.
Уход за LM-активатором
- после каждого использования lm-активатор желательно промывать в проточной теплой воде;
- для тщательной очистки lm-активатора желательно дважды в неделю применять растворимые таблетки Protefix
- скученность зубов в области резцов и клыков;
- ротация резцов и клыков;
- детализация и положения зубов после лечения брекет-системой;
- десневая улыбка;
- открытый прикус;
- дистальный прикус;
- перекрестный прикус
- мезиальный прикус;
- смещение серединной линии, превышающее 3 мм;
- очень узкая верхняя зубная дуга.
 Доступны два варианта низкой модели – Низкая короткая (LOW) и Низкая длинная (LOW long). Низкая короткая модель – модель, укороченная в области моляров, для пациентов, у которых вторые моляры еще не прорезались (желтый контейнер). Низкая длинная модель – модель с удлиненной областью моляров, для пациентов, у которых уже прорезались вторые моляры (синий контейнер).
Доступны два варианта низкой модели – Низкая короткая (LOW) и Низкая длинная (LOW long). Низкая короткая модель – модель, укороченная в области моляров, для пациентов, у которых вторые моляры еще не прорезались (желтый контейнер). Низкая длинная модель – модель с удлиненной областью моляров, для пациентов, у которых уже прорезались вторые моляры (синий контейнер).Мы являемся официальным магазином LM-активаторов в России. У нас можно купить 100%-оригинальные LM-активаторы всех видов и моделей. Быстрая доставка по всей России. Заказывайте:
Быстрая доставка по всей России. Заказывайте:
LM-Активаторы в практике врача-ортодонта — Рауденталл
Ортодонтическое лечение пациентов с патологией зубочелюстной системы – это неотъемлемая часть комплексного стоматологического лечения. Такие слова как «пластинки», «брекеты» вошли в наш лексикон наравне с такими стоматологическими понятиями, как «пломба», «коронка», «зубной протез». Уже трудно встретить подростка, не носившего в своей жизни брекеты. Уже все чаще родители настаивают на ортодонтическом лечении для своих детей в раннем дошкольном возрасте. Уже не редко на консультацию врача-ортодонта приносят детей при прорезывании первой пары зубов и наличии диастемы.
Эффективно лечить различные патологии постоянного прикуса ортодонты научились. В их распоряжении всевозможные съемные аппараты, брекет — системы, челюстно — лицевая хирургия. Подростки «плачут» и носят все необходимое для создания социально приемлемой картины успешного человека, а взрослые все чаще решаются на хирургическое изменение прикуса и лица для достижения той же самой цели.
Речь идет о ПРОФИЛАКТИКЕ зубочелюстных аномалий и РАННЕМ ЛЕЧЕНИИ первых проявлений отклонения от нормы. Здесь важен ликбез родителей о необходимости грудного вскармливания, своевременного отказа от соски, борьбе с вредными привычками сосания пальца, ротового дыхания и т. д. Очень важно и скрининговое обследование максимального количества детей у врача-ортодонта. Важно своевременное лечение выявленных отклонений зубочелюстной системы и систем, напрямую влияющих на формирование прикуса, здоровье ЛОР органов, позвоночника от нормы.
Важно своевременное лечение выявленных отклонений зубочелюстной системы и систем, напрямую влияющих на формирование прикуса, здоровье ЛОР органов, позвоночника от нормы.
К сожалению, система диспансеризации населения была разрушена в печальные 90-е годы прошлого века, закрылись «зубные кабинеты» в школах, прекратились плановые осмотры детей в детских садах, а частная стоматология не делает ставку на прием детей вследствие малого финансового результата по сравнению с протезированием на имплантатах. И напрасно.
На детском приеме в ОРТОНИКЕ каждый ребенок осматривается «глазами трех специалистов»: терапевта, хирурга, ортодонта. Естественно, в первую очередь, оценивается и приводится в норму уровень гигиены полости рта. Затем начинается терапевтическая и хирургическая санация полости рта. Дети, нуждающиеся в помощи врача ортодонта, проходят дополнительное обследование: диагностические фото, контрольно- диагностические модели челюстей, рентгеновские снимки.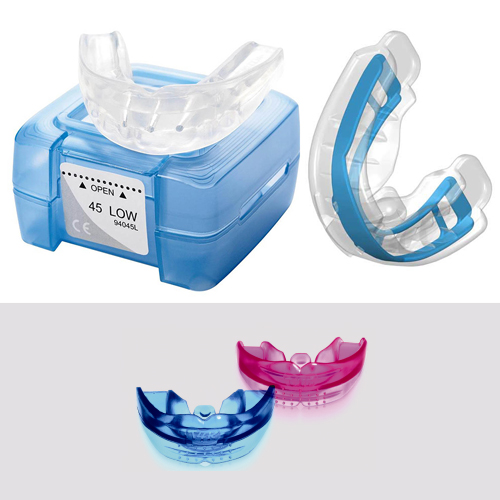
В ортодонтическом лечении пациентов ОРТОНИКИ мы используем съемные пластинки, моно- и твинблоковые активаторы, и конечно же, силиконовые активаторы.
Из множества имеющихся на рынке активаторов мы выбрали LM-активатор финского производства. На этот выбор повлияло обучение у автора LM-технологии профессора Катри Кески-Нисула в январе 2013 года в Университете г. Турку. А результаты, полученные при лечении наших пациентов за 2013 год, утвердили нас в правильности этого выбора.
LM-активатор удобен и эффективен в лечении многих типов малокклюзии, в том числе скученности, открытого, глубокого и дистального прикуса. Линейка размеров аппарата от 10 до 70 позволяет подобрать аппарат для детей разного возраста с большой точностью. К тому же LM-активатор требует лишь минимальной корректировки. Время, проводимое пациентом в кресле составляет от 5 до 12 минут. Кроме того, лечение данным аппаратом увеличивает интервалы между посещениями до 6-12 недель. Также этот же аппарат может быть использован и в качестве ретейнера. Существующие модели LM-активатора не эффективны для пациентов со скелетной формой 3 класса, и поэтому не назначались для них.
К тому же LM-активатор требует лишь минимальной корректировки. Время, проводимое пациентом в кресле составляет от 5 до 12 минут. Кроме того, лечение данным аппаратом увеличивает интервалы между посещениями до 6-12 недель. Также этот же аппарат может быть использован и в качестве ретейнера. Существующие модели LM-активатора не эффективны для пациентов со скелетной формой 3 класса, и поэтому не назначались для них.
МАТЕРИАЛЫ
За 2013 год назначение использовать в лечении LM-активатор получили 102 пациента разного возраста. Из них 2 ребенка с молочным прикусом, 84 ребенка со сменным и 2 взрослых пациента с постоянным прикусом. Лечение начиналось после клинического осмотра, анализа контрольно-диагностических моделей, фоторегистрации лица и зубных рядов. Рентгеновские снимки: ОПТГ назначался всем пациентам с 7 лет, ТРГ- при выраженных дистальном или мезиальном прикусе. Один или два LM-активатора были использованы у каждого пациента. Смена аппарата требовалась при необходимости перейти на следующий размер или при деформации аппарата вследствие «противоправных» действий маленького пациента. Размер аппарата определялся в соответствии с рекомендациями завода-изготовителя. Пациенты получали рекомендации носить аппарата всю ночь и 2 часа днем. Фактически, пациенты аппарат носили только ночью. Единичные дисциплинированные пациенты носили аппарат дополнительно один час днем.
Смена аппарата требовалась при необходимости перейти на следующий размер или при деформации аппарата вследствие «противоправных» действий маленького пациента. Размер аппарата определялся в соответствии с рекомендациями завода-изготовителя. Пациенты получали рекомендации носить аппарата всю ночь и 2 часа днем. Фактически, пациенты аппарат носили только ночью. Единичные дисциплинированные пациенты носили аппарат дополнительно один час днем.
Продолжительность наблюдения группы пациентов с января 2013 года составляет 12 месяцев.
Примеры лечения:
Л.А. возраст 10/3. Дистальная окклюзия, глубокое резцовое перекрытие,скученное положение зубов. Модель LM-активатора — №45 Low Short. Устранение протрузии 12, 22, ротации 11,21, начальное выравнивание нижних резцов при использовании аппарата только в ночное время на протяжении 12 месяцев 2013 года.
Рис.1-3 Дата: 13. 01.2013
01.2013
Рис.4-6 Дата: 12.2013
Н.В. возраст 11/6. Автомобильная травма. Полный вывих 11, вколоченный вывих 21 22. Модель LM-активатора — №50 Low Short. Экструзия 21 22 эластичными кольцами с опорой на крючок в базисе аппарата. Аппарат использовался параллельно со съемными протезами на верхнюю и нижнюю челюсть в течение четырех месяцев.
Даты: 11.09.2012 — 18.01.2013 — 05.08.2013 — 26.12.2013
Ч.А. возраст 22/3. Модель LM-активатора — №60 Low Long использовался в качестве ретенционного аппарата после ортодонтического лечения.
ОБСУЖДЕНИЕ
Наша статистика показала, что на адаптацию к аппарату требовалось от нескольких дней до 3-4 недель. Двум пациентам потребовалась дополнительная коррекция аппарата в подъязычной области, остальные прекратили предъявлять жалобы на дискомфорт от аппарата на 3-4 день ежедневного использования. К концу первого месяца лечения «находить» аппарат на подушке прекратили все пациенты, кроме 5 пациентов из 102. Эти пациенты прекратили лечение. Первые зарегистрированные с помощью фото улучшения положения зубов были достигнуты уже через 4-6 недель. Прием пациентов производился каждые 6-8 недель. Пациент рассказывал о своих впечатлениях об аппарате. На каждом последующем осмотре отмечались позитивные перемены. В течение года наблюдения большинство детей, получавших лечение, показали благоприятные межчелюстные соотношения в сегменте резцов, клыков и моляров. Закрывалась вертикальная щель во фронтальном отделе при открытом прикусе, уменьшалась глубина резцового перекрытия при глубоком прикусе, выравнивались зубных рядов, появлялся мезиальный шаг в области моляров, а клыки показывали почти полный I класс. И все это при использовании аппарата практически у 99% детей лишь в ночное время.
К концу первого месяца лечения «находить» аппарат на подушке прекратили все пациенты, кроме 5 пациентов из 102. Эти пациенты прекратили лечение. Первые зарегистрированные с помощью фото улучшения положения зубов были достигнуты уже через 4-6 недель. Прием пациентов производился каждые 6-8 недель. Пациент рассказывал о своих впечатлениях об аппарате. На каждом последующем осмотре отмечались позитивные перемены. В течение года наблюдения большинство детей, получавших лечение, показали благоприятные межчелюстные соотношения в сегменте резцов, клыков и моляров. Закрывалась вертикальная щель во фронтальном отделе при открытом прикусе, уменьшалась глубина резцового перекрытия при глубоком прикусе, выравнивались зубных рядов, появлялся мезиальный шаг в области моляров, а клыки показывали почти полный I класс. И все это при использовании аппарата практически у 99% детей лишь в ночное время.
Время нахождения пациента в клинике, потребовавшееся для всего курса лечения, было значительно меньше, чем у других техник, потому как регулярные посещения каждые 4-6 недель занимали всего 5-10 минут каждое. Преимущество LM- активатора в том, что он не только направляет прорезывание зубов, но и одновременно действует на зубные дуги в трансверзиальном и сагиттальном направлениях.
Преимущество LM- активатора в том, что он не только направляет прорезывание зубов, но и одновременно действует на зубные дуги в трансверзиальном и сагиттальном направлениях.
Родительское воспитание и поддержка всегда необходимы при лечении детей с применением съемных аппаратов, особенно в начале лечения. Большинство детей быстро освоили аппарат, от родителей 8 пациентов за год мы получили нарекания из-за многократных выпадений аппарата изо рта ребенка во время ночного сна. У 7 детей выпадение аппарата изо рта прекратилось через 1-1,5 месяца использования. Одного ребенка мы не смогли вызвать на прием по причине отказа родителей. Прекращение лечения могло быть связано с проблемами при соблюдении режима ношения, либо ребенок не хотел носить устройство, либо его родители не были мотивированы достаточно, чтобы поддержать его. Различные психосоциальные нарушения в семьях этих детей были одной из главных причин плохого сотрудничества. В материалах профессора Катри Кески-Нисула было высказано предположение, что молодой возраст пациентов будет основным ограничивающим фактором для широкого распространения раннего лечения. В нашей практике оказалось, что самым важным фактором, влияющим на соблюдение режима ношения, является семейное положение, а не возраст самих пациентов. На основании статистики ОРТОНИКИ мы не можем делать глобальных выводов эффективности лечения в раннем смешанном прикусе, но мы будем наблюдать наших пациентов дальше. Однако полученные результаты уже радуют. Есть и результаты, которых мы не ожидали: гигинена полости рта у пациентов, носящих LM- активатор и проходящих регулярные осмотры лучше, таковой у пациентов, не нуждающихся в ортодонтическом лечении и проходящих осмотры лишь 1 раз в 6 месяцев. Прирост кариеса и прирост осложненных форм кариеса у этих пациентов ниже на 16 %. Маленькие пациенты, привыкая к манипуляциям врача, легко воспринимают и другие стоматологические процедуры: профгигиену, лечение зубов. Возросли и загрузка клиники на профилактическом приеме и на приеме детского стоматолога, улучшились и финансовые результаты, что немаловажно для частной клиники.
В нашей практике оказалось, что самым важным фактором, влияющим на соблюдение режима ношения, является семейное положение, а не возраст самих пациентов. На основании статистики ОРТОНИКИ мы не можем делать глобальных выводов эффективности лечения в раннем смешанном прикусе, но мы будем наблюдать наших пациентов дальше. Однако полученные результаты уже радуют. Есть и результаты, которых мы не ожидали: гигинена полости рта у пациентов, носящих LM- активатор и проходящих регулярные осмотры лучше, таковой у пациентов, не нуждающихся в ортодонтическом лечении и проходящих осмотры лишь 1 раз в 6 месяцев. Прирост кариеса и прирост осложненных форм кариеса у этих пациентов ниже на 16 %. Маленькие пациенты, привыкая к манипуляциям врача, легко воспринимают и другие стоматологические процедуры: профгигиену, лечение зубов. Возросли и загрузка клиники на профилактическом приеме и на приеме детского стоматолога, улучшились и финансовые результаты, что немаловажно для частной клиники.
Выводы:
Использование LM-активаторов в практике врача ортодонта на детском приеме, по результатам лечения ортодонтических пациентов, эффективно. Финансовые показатели работы частной стоматологической клиники при использовании LM-активаторов позитивные. Мы рекомендуем использовать LM-активатор в практике врача стоматолога детского и врача ортодонта.
Финансовые показатели работы частной стоматологической клиники при использовании LM-активаторов позитивные. Мы рекомендуем использовать LM-активатор в практике врача стоматолога детского и врача ортодонта.
Литература:
1 ПерсинЛ.С. «Ортодонтия. Диагностика и виды зубочелюмтных аномалий.»
2 W.Proffit Contemporary orthodontics.
Автор статьи:
Н.М. Соснина, врач-ортодонт. Семейная стоматология «ОРТОНИКА» Тольятти, ortonika.ru
LM-активаторы
LM-активаторы. Ортодонтическая коррекция сменного или раннего постоянного прикуса
LM-активаторы — ортодонтические конструкции на основе медицинского силикона. Используются в преимущественно у детей и подростков. Хорошие результаты достигаются в молочном, смешанном и начальном постоянном прикусе прикусе.
Использование LM-активаторов способствует
- правильному развитию челюстей
- появлению постоянных зубов
- развитию контактов зубов нижней и верхней челюстей
- правильному расположению языка
- улучшению дикции
Терапия LM-активаторами позволяет обойтись без сложного ортодонтического лечения. При точном соблюдении рекомендаций врача-ортодонта меры терапии очень эффективны.
При точном соблюдении рекомендаций врача-ортодонта меры терапии очень эффективны.
Конструкция LM-активатора
LM-активаторы делаются из гипоаллергеного силиконового материала. Визуально устройство напоминает каппу, используемую боксерами. Материал выбран удачно, пациенты быстро привыкают к LM-активатору.
Форма активаторов идентична правильному прикусу. Большой выбор моделей позволяет подобрать подходящий аппарат для каждого конкретного пациента. Если ребенок испытывает при ношении неудобства, например сильное давление, действие устройства можно изменить.
Конструкция двухчелюстная. Встречаются высокие, низкие, длинные и короткие типы LM-активаторов.
- Высокие короткие модели расширены в области вторых премоляров и моляров. Исправляют скелетные и зубочелюстные аномалий при открытом прикусе;
- Низкие длинные модели удлинены в области моляров. Необходимы детям с прорезавшимися вторыми молярами.
Тип и размер LM-активатора подбирают индивидуально, учитывая размеры передних зубов и используя линейку LM-OrthoSizer.
В передней части LM-активаторов находятся дыхательные отверстия. Они необходимы пациентам, имеющим проблемы с носовым дыханием. Правильное положение передних зубов определяется выемками. Активатор носится с закрытым ртом и сомкнутыми зубами.
LM-активаторы
- исправляют прикус и выравнивают зубы
- устанавливают челюсти в правильное соотношение
- нормализуют дыхательные процессы
- облегчают глотание
- нормализуют тонус мышц лица и челюстей
- помогают избавиться от вредных привычек вроде грызения ногтей и сосания пальцев
- исключают вероятность будущих рецидивов
- служат для профилактики ортодонтических аномалий
Области использования
Показания для использования LM-активаторов
- дистальный, глубокий, открытый, перекрёстный типы прикуса
- профилактика аномалий прикуса
- тесное положение фронтальной группы зубов
- обратное положение отдельных зубов
- десневая улыбка
Противопоказания
- антериальный прикус
- смещение средней линии более, чем на три миллиметра
- зауженный ряд верхних зубов
LM-активатор у взрослых пациентов
Активаторы применяют у взрослых
- при патологической стираемости эмали и дисфункции височно-нижнечелюстных суставов
- как каппу для реминерализующей терапии
- как ретейннер после ортодонтического лечения
Врачи-ортодонты постоянно находят новые области использования для LM-активаторов.
Достоинства
- успешно исправляют прикус и дикцию
- удобны и просты в использовании
- более эстетичны в сравнении с брекетами
- тонизируют круговую мышцу рта
- помогают отказаться от вредных привычек
- не вызывают аллергии
- стоят дешевле брекет-систем
Недостатки
- иногда выпадают из ротовой полости во время сна
Уход
За LM-активаторами можно ухаживать самостоятельно. Простой вид обслуживания — промывание конструкции в проточной воде. Достаточно щетки и зубной пасты. Футляр можно мыть в посудомоечной машине.
Продолжительность лечения
Сроки ношения LM-активатора определяются возрастом пациента, особенностями организма и выраженностью патологии. В среднем требуется не менее года. Может возникнуть необходимость в нескольких активаторах из-за взросления детей.
Особенности ношения
Наиболее благоприятное время для ношения LM-активатора — ночь. Днем можно его одевать часа на два. В зависимости от конкретной ситуации можно обойтись только ночным ношением или наоборот, требуется продолжительное дневное. Сроки ношения аппарата ночью и днем определяет врач-ортодонт.
Днем можно его одевать часа на два. В зависимости от конкретной ситуации можно обойтись только ночным ношением или наоборот, требуется продолжительное дневное. Сроки ношения аппарата ночью и днем определяет врач-ортодонт.
При ношении LM-активатора необходимо правильно зафиксировать зубы и сжать губы. Аппарат нельзя прикусывать или жевать, чтобы не повредить его.
Резюме
LM-активаторы — особые ортодонтические конструкции для коррекции прикуса, безопасные, эффективные и удобные. Их можно носить всего несколько часов днем. Необходим ли лично Вам LM-активатор — ответит только профессиональный ортодонт. Мы приглашаем Вас в клинику стоматологи «Ювелирная работа» на консультации и лечение.
ЛМ-Активатор, аппарат для раннего ортодонтического лечения
Петрова Ольга (мама)
Добрый день!
Хочу оставить свой отзыв в помощь родителям, которые столкнулись с проблемой ортодонтии у своих детей и не знают куда и к какому врачу обратиться!
Сама столкнулась с такой проблемой около 2-х лет назад, когда искала куда обратиться с сыном -подростком для консультации и дальнейшего лечения. Ходили в городскую пол-ку, показывали разным врачам, но кроме страха и не желания что-то исправлять у сына не бы ло ничего!!!
Ходили в городскую пол-ку, показывали разным врачам, но кроме страха и не желания что-то исправлять у сына не бы ло ничего!!!
Случайно, в интернете, наткнулась на Вашу сеть клиник, нашла ближайшую к дому пол-ку, м.»Тульская», записались на консультацию к доктору Блиндер Ж.А., для себя решив, что последний раз сходим, проконсультируемся и все….НО Жанна Аркадьевна с первого нашего посещения нашла подход к сыну-подростку!!!!, смогла его убедить в необходимости лечения и установке брекет-системы!!!!
Жанна Аркадьевна, спасибо Вам огромное за Ваш профессионализм, за кропотливый труд, на протяжении всего времени лечения (1.5 года!!), за т от позитив, которым Вы заряжаете всех своих пациентов!!!! и конечно же- за результат!!!, который был достигнут, благодаря работе всей Вашей команды!!! Отдельное спасибо гигиенисту Галине Владимировне, за ее помощь, наставничество и ТЕРПЕНИЕ!!!! Вашему ассистенту- Екатерине!
Вашим замечательным администраторам, Екатерине и Виктории, которые на протяжении всего лечения очень тепло встречали нас и всех Ваших пациентов. В настоящее время, по нашей рекомендации у Жанны Аркадьевны лечатся дети-подростки, наших знакомых , лечение проходит замечательно!!! дети и родители -довольны!!!! Желаем им скорейшего окончания лечения и таких же великолепных результатов!
В настоящее время, по нашей рекомендации у Жанны Аркадьевны лечатся дети-подростки, наших знакомых , лечение проходит замечательно!!! дети и родители -довольны!!!! Желаем им скорейшего окончания лечения и таких же великолепных результатов!
Уважаемые родители, мамы, если Вы ищите лучшую клинику и врача с «золотыми руками» -Вам в «Ортодонт-центр» на м.»Тульская», к врачу БЛИНДЕР ЖАННЕ АРКАДЬЕВНЕ! С ней Ваши дети будут в надежных руках!, а результат не заставит себя ждать! С уважением, Петрова Ольга
 
 
LM-активатор для зубов в СПб, цены
Показанием к применению являются нарушение прикуса (дистальный, глубокий, открытый, перекрестный), профилактика прикуса, тесное положение передней группы зубов, обратное положение некоторых зубов, а также «десневая» улыбка. Наибольшего эффекта трейнер дает при молочном и смешанном прикусе. При начальном постоянном прикусе аппарат тоже показывает хорошие результаты, а вот взрослым его редко назначают. Например, у взрослых ЛМ-активатор применяют при дисфункции височно-нижнечелюстных суставов или при высокой стираемости твердых тканей зубов. Иногда его используют как каппу для реминерализирующей терапии или ретенционную конструкцию после ортодонтического лечения.
Например, у взрослых ЛМ-активатор применяют при дисфункции височно-нижнечелюстных суставов или при высокой стираемости твердых тканей зубов. Иногда его используют как каппу для реминерализирующей терапии или ретенционную конструкцию после ортодонтического лечения.
Трейнер изготавливается из очень эстетичного медицинского силикона, который позволяет пациенту быстро привыкнуть к аппарату. ЛМ-активатор предназначен как для верхней, так и для нижней челюстей. Он имеет достаточно высокие стенки и дыхательные отверстия, расположенные в передней части.
В процессе ношения аппарата зубы должны быть максимально сомкнуты, а рот закрыт, поэтому для передних зубов есть специальные выемки, позволяющие им принимать правильное положение.
Противопоказания к применению аппарата
Аппарат не применяется при мезиальном прикусе, смещении средней линии на 3 и более мм, очень узком верхнем зубном ряде.
Модели ЛМ-активаторов
Выпускаются высокая и низкая модели аппарата. Первая имеет утолщение силикона в боковых участках и применяется для лечения открытого прикуса. Низкая модель имеет более широкий спектр назначения.
Низкая модель имеет более широкий спектр назначения.
Виды ЛМ-активаторов
Все модели различаются по внешнему виду. Есть короткие и длинные виды. Короткие ЛМ-активаторы представляют собой конструкции, укороченные в задних отделах, которые применяются до прорезывания седьмых зубов.
Все модели ЛМ-активаторов имеют разные размеры. Для каждого пациента размер подбирается индивидуально, с помощью специальной линейки или расчетов.
Сроки лечения
Длительность лечения всегда индивидуальна и будет зависеть от возраста пациента, вида и степени патологии. Зачастую лечение трейнером занимает меньше года.
Носить этот аппарат рекомендуют ночью и днем в течение двух часов. Врач после осмотра пациента может сказать, что ночного ношения будет достаточно, либо может увеличить еще и дневное ношение.
Кукшина Лилия Сергеевна
02 Июня 2021
Добрый день! Вчера 01.06.2021 мы лечили зубки во сне. Хочу выразить огромную благодарность всей команде, которая с нами работала, они просто мега супер специалисты. Все прошло хорошо, ребёнок перенёс все хорошо. Радуется, что зубки вылечили, а что фея пришла и забрала зубки, не очень рада, хотя раньше все ждала, когда же у неё выпадут зубки для феи))) Осадчая Алла Павловна с первых минут завоевала доверие у моих детей, да и я смогла ей довериться и принять очень тяжелое для меня решение — лечить во сне. Спасибо большое. Мы теперь будем ходить только к вам))) Наш анестезиолог Моисеенко Кирилл Александрович, все рассказал, я доверила ему ребёнка, и вы знаете, он — молодец. Советую, очень хороший специалист. Медперсонал очень крутой!!! Большое спасибо вам!!!
Все прошло хорошо, ребёнок перенёс все хорошо. Радуется, что зубки вылечили, а что фея пришла и забрала зубки, не очень рада, хотя раньше все ждала, когда же у неё выпадут зубки для феи))) Осадчая Алла Павловна с первых минут завоевала доверие у моих детей, да и я смогла ей довериться и принять очень тяжелое для меня решение — лечить во сне. Спасибо большое. Мы теперь будем ходить только к вам))) Наш анестезиолог Моисеенко Кирилл Александрович, все рассказал, я доверила ему ребёнка, и вы знаете, он — молодец. Советую, очень хороший специалист. Медперсонал очень крутой!!! Большое спасибо вам!!!
Гнедькова Олеся Александровна
15 Мая 2021
Выражаю огромную благодарность Лобко Татьяне Владимировне и её ассистентке Лиане. Татьяна Владимировна настоящий профессионал своего дела. Так виртуозно , быстро и качественно делает свою работу. Я под большим впечатлением! Берегите её)!
читать далее…Костюк Людмила Евгеньевна
13 Мая 2021
Выражаю ОГРОМНЕЙШУЮ благодарность доктору Похил Виктории Игоревне! Не могу даже представить более профессионального и доброжелательного подхода к ребёнку!!! На каждый приём дочь идёт с удовольствием!! Спасибо большое за наши зубки!!!
читать далее. ..
..Галина
10 Мая 2021
У ребёнка, к сожалению, очень рано развился кариес, прогрессировал. Были в 5 популярных клиниках города, но решили довериться клинике «Вероника», доктору Гай Екатерине Михайловне. Очень остались довольны, лечение было под медикаментозным сном, все прошло хорошо. Хочется выразить огромную благодарность всему персоналу, низкий до Земли поклон анестезиологу Козыренко Антону Сергеевичу, Екатерине Михайловне, администраторам за тёплый приём и встречу — огромнейшее спасибо.
читать далее…Авдеева Мария Владимировна
14 Апреля 2021
Выражаем большую благодарность Гай Екатерине Михайловне и Кособрюхову Михаилу Юрьевичу. Врачи с большой буквы и огромным сердцем. Спасибо Вам за красивую улыбку моего сына.
читать далее…Мария
11 Апреля 2021
Хочу выразить благодарность и сказать огромное спвсибо Журавлевой Ирине Сергеевна за огромный профессионализм и чуткое отношение!Приехали с острой болью.
Она мягко находит подход к детям, доброжелательная!
Перед первым приемом немного волновались, что не захотят открыть рот, чтобы показать зубки, будут плакать, но Ирина Сергеевна в игровой в форме справилась. Мы были удивлены, так как уже был опыт с походом в другие клиники и там заканчивалось печально.
Сделали снимок и решили удалить зуб. Все прошло идеально.
Роману посещение стоматолога понравилось.
В самой клинике обстановка приятная, все врачи доброжелательные, и персонал приветливый.
Однозначно рекомендуем всем!
LM-Activator — Медицинская техника
ПРОДУКЦИЯ LM-ACTIVATOR
| Миофункциональные аппараты | |
Миофункциональный аппарат LM-Activator сделан из биосовместимого и эластичного силикона, делающего его гигиеничным и удобным в ношении. LM — Activator имеет дополнительные отверстия во фронтальной части, облегчающие дыхание и делающее возможным ношение аппарата при заболеваниях ЛОР-органов.Силикон достаточно мягкий, чтобы обеспечить удобство ношения и в то же время достаточно жесткий, чтобы пациент не мог повредить его.В домашних условиях трейнер чистится зубной щеткой с пастой, дезинфицируется кипячением, в условиях стоматологической клиники трейнер автоклавируется.Имеется две модификации трейнеров — низкая модель, для коррекции глубокого прикуса и высокая, для лечения открытого прикуса. Существует 13 размеров низкой модели и 11 — высокой.Для удобства определения размеров используется специальная линейка (LM-OrthoSizer). LM — Activator имеет дополнительные отверстия во фронтальной части, облегчающие дыхание и делающее возможным ношение аппарата при заболеваниях ЛОР-органов.Силикон достаточно мягкий, чтобы обеспечить удобство ношения и в то же время достаточно жесткий, чтобы пациент не мог повредить его.В домашних условиях трейнер чистится зубной щеткой с пастой, дезинфицируется кипячением, в условиях стоматологической клиники трейнер автоклавируется.Имеется две модификации трейнеров — низкая модель, для коррекции глубокого прикуса и высокая, для лечения открытого прикуса. Существует 13 размеров низкой модели и 11 — высокой.Для удобства определения размеров используется специальная линейка (LM-OrthoSizer).Показаниями для лечения с помощью LM-Activatorа являются: -профилактика развития аномалий прикуса за счет устранения вредных привычек и восстановления правильных функций зубочелюстной системы;-нарушение прикуса в горизонтальной (дистальный прикус), вертикальной (открытый и глубокий прикус) и трансверзальной (перекрестный прикус) плоскостях;-скученность зубов в области резцов и клыков и их ротация;-«десневая» улыбка возможно использование LM-Activator’а в качестве детализатора и ретенционного аппарата, после лечения с помощью брекет-системы.  |
|

Каждый год мы оплачиваем 20-ти лучшим ортодонтам круиз с компанией ЛМ-Активатор. На круизном лайнере по маршруту Турку-Стокгольм доктора обучаются и наслаждаются поездкой. Надеемся увидеть Вас в следующем круизе!
Мастер-класс ЛМ Активатор, Финляндия, февраль 2014год
LM-активаторы (трейнера) в Минске: цены, отзывы
LM-activator намного удобнее современных брекет-систем, поскольку не требует постоянного ношения. Чтобы решить проблемы с зубами, достаточно носить его на протяжении 12 часов в день. Изготовляется конструкция по слепкам зубов пациента.
Эффективность LM-активатора
LM-активатор, помимо пластмассовой пластины, может состоять из пружин, петель, дужек и специальных винтов. Его конструкция устанавливается на поверхность зубов изнутри и слизистую оболочку твёрдого нёба и способна смещаться вперёд, назад, в стороны или сразу в нескольких направлениях. Пластмассовая пластина выпиливается в том направлении, куда зуб должен переместиться.
Пластмассовая пластина выпиливается в том направлении, куда зуб должен переместиться.
LM-активатор устанавливают для следующих целей:
- для фиксации правильного положения челюсти;
- для обеспечения правильного прорезания постоянных зубов;
- для выпрямления зубов искривлённых;
- для избавления от неправильного прикуса;
- для постановки языка в правильное положение;
- для выдвижения нижней челюсти;
- для лечения скученности зубов.
Надевать ЛМ-активатор неприятно только в первый раз. Однако он эффективен для исправления любых ортодонтических нарушений уже в детские годы. К тому же можно облегчить ношение такой конструкции, сделав в пластине специальные отверстия, что важно для пациентов с ЛОР-заболеваниями или затруднённым носовым дыханием.
После установки конструкции рекомендуется почаще посещать центр стоматологии, чтобы врач мог отслеживать ход лечения. Возможно, спустя определённый период трейнер придётся сменить.
Виды активаторов
- Андрезена-Гойпля. Этот LM-activator представляет собой многоблоковую конструкцию, фиксирующую нижнюю челюсть и лечащую скученность зубов. При необходимости расширения зубов в конструкцию вставляют пружину Коффина. Данный LM-activator позволяет перестраивать соотношение рядов зубов. Используется он не только для избавления от ортодонтических аномалий, но и для исправления дистального прикуса;
- LM. Этот LM-activator представляет собой гипоаллергенную силиконовую конструкцию, удерживающую зубы в нужном положении. Она необходима для лечения аномального прорезывания и скученности коренных зубов. Конструкция должна надеваться на ночь. Днём же её можно установить лишь на два часа. Затем её ополаскивают и кладут в специальный контейнер. Активатор предназначен для детей в возрасте 3-12 лет. ЛМ-активаторы делятся на два типа: призванные лечить патологии и призванные лечить открытый прикус. Названные выше их разновидности не являются единственными.
 Перечень здесь достаточно широк, благодаря чему в каждом конкретном случае используется индивидуальная конструкция.
Перечень здесь достаточно широк, благодаря чему в каждом конкретном случае используется индивидуальная конструкция.
 Перечень здесь достаточно широк, благодаря чему в каждом конкретном случае используется индивидуальная конструкция.
Перечень здесь достаточно широк, благодаря чему в каждом конкретном случае используется индивидуальная конструкция.Следует помнить, что для правильного прикуса недостаточно установить ЛМ-активатор. Важное значение имеет формирование у ребёнка привычки ухаживать за зубами. Так, привычку их чистить можно формировать, начиная с 5 лет – заодно это превосходная профилактика кариеса. Полезны тренировки зубо-челюстного аппарата, когда ребёнку даётся грубая пища. Сладкое же следует ограничить, а после его употребления – полоскать рот. Если соблюдать данные рекомендации, то платные услуги стоматолога во многих случаях не понадобятся.
К мерам профилактики проблем с прикусом относится и своевременное лечение ЛОР-инфекций, дабы не возникали проблемы с дыханием через нос, а также обращение к стоматологу за профессиональной чисткой зубов, начиная с четырёхлетнего возраста.
Получаем красивую улыбку с детства
Чтобы улыбка ребёнка была красивой, необходимо вовремя предотвращать проблемы с зубами. В клинку обращаются все желающие установить трейнер для зубов.
В клинку обращаются все желающие установить трейнер для зубов.
Ортодонты «Аладен» имеют опыт в проведении сложнейших операций и пользуются качественными материалами от зарекомендовавших себя поставщиков. Обратившись в клинику, можно быть уверенным в том, что высокое качество здесь гарантируется. В клинике цена на трейнер достаточно приемлема.
LM-Activator ™ — LM-Dental
Силиконовый активатор и выравниватель
LM-Activator ™ — это активатор и выравниватель анатомической формы для ортодонтического лечения. Он выравнивает зубы и активирует рост нижней челюсти (то есть выдвигает нижнюю челюсть вперед для коррекции класса II) и может одновременно увеличивать длину зубной дуги, чтобы исправить или избежать скученности. LM-Activator ™ бережно поддерживает здоровый рост зубов и челюстей.
«Одним из замечательных свойств устройства является то, что он может устранять несколько проблем одновременно.Большинство других ортодонтических аппаратов сосредоточены на решении одной проблемы за раз ».
— профессор Юха Варрела, Университет Турку, Финляндия

LM-Activator ™ эффективно корректирует различные аномалии прикуса; чрезмерный прикус, глубокий прикус, открытый прикус, скученность, класс II и передний перекрестный прикус зубов, то есть перевернутые резцы. Корректировать сагиттальные и вертикальные взаимоотношения можно одновременно.
«Мы можем контролировать окклюзию в трехмерном пространстве с помощью LM-Activator ™. Мы можем контролировать чрезмерный прикус, чрезмерный прикус, боковые контакты нижней и верхней дуги и изменять сагиттальный рост.”
— адъюнкт-профессор Джоаккино Пеллегрино, Неаполитанский университет имени Федерико II, Италия
LM-Activator ™ является съемным и в основном используется в ночное время. Он изготовлен из гибкого и удобного прозрачного медицинского силикона, полностью биосовместим и не содержит добавок.
«Когда я увидел результаты LM-Activator ™, я был удивлен.
— Профессор Юха Варрела, Университет Турку, Финляндия
LM-Activator ™
LM-Activator ™ — это первое поколение LM-Activator ™, представленное в 2004 году.Он помог сотням тысяч детей во всем мире обрести здоровую улыбку.
LM-Activator ™ доступен в следующих моделях:
- Короткий: короткие модели имеют более короткое молярное сечение. Для пациентов, у которых вторые моляры еще не прорезались.
- Длинный: у длинных моделей более длинное молярное сечение. Для пациентов, у которых прорезались вторые моляры.
- Low: модели Low могут применяться во многих различных случаях.
- Высокий: Специально разработан для лечения открытых прикусов скелетных и зубочелюстных суставов.Толще в области вторых премоляров и коренных зубов.
Узкая модель LM-Activator ™ 2 сравнима с оригинальной LM-Activator ™. Мы рекомендуем использовать LM-Activator ™ 2 из-за его улучшенных характеристик и функциональности в материалах и дизайне.
Эпигенетическая и традиционная регуляция распределена между активаторами FLO11, позволяя настраивать неоднородность на уровне популяции в ее экспрессии
Abstract
Эпигенетические переключатели кодируют информацию о своем состоянии либо локально, часто посредством ковалентной модификации ДНК или гистонов, либо глобально, обычно на уровне регулирующего фактора trans .Здесь мы исследуем, как регуляция эпигенетических переключений, кодируемых цис , контролирует степень гетерогенности экспрессии генов, которая в конечном итоге связана с фенотипическим разнообразием в популяции. Мы показываем, что две копии локуса FLO11 в Saccharomyces cerevisiae переключаются между молчащим и компетентным состоянием промотора случайным и независимым образом, подразумевая, что молекулярное событие, приводящее к переходу, происходит локально на промоторе, в цис .Мы далее количественно оцениваем влияние регуляторов trans как на медленные эпигенетические переходы между молчащим и компетентным состоянием промотора, так и на быстрые переходы промотора, связанные с традиционной регуляцией FLO11 . Мы обнаружили, что разные классы регуляторов влияют на эпигенетическую, общепринятую или обе формы регуляции. Распределение кинетического контроля эпигенетического молчания и обычной активации генов предлагает клеткам гибкость в формировании распределения экспрессии генов и фенотипа в популяции.
Мы обнаружили, что разные классы регуляторов влияют на эпигенетическую, общепринятую или обе формы регуляции. Распределение кинетического контроля эпигенетического молчания и обычной активации генов предлагает клеткам гибкость в формировании распределения экспрессии генов и фенотипа в популяции.
Сведения об авторе
В нестабильном и меняющемся мире микробные популяции с разнообразным диапазоном фенотипов могут превосходить монолитную популяцию. На протяжении многих поколений мутации могут приводить к генетическому разнообразию в популяции. Однако у микробов есть стратегии для быстрого создания такого разнообразия. Например, если несколько генов включаются и выключаются медленно, случайным образом и независимо друг от друга, то возможна большая комбинация состояний экспрессии генов и, следовательно, фенотипов.Различные состояния экспрессии генов не связаны с изменениями в последовательности ДНК и, следовательно, наследуются эпигенетически. Мы показываем, что две копии гена FLO11 в S. cerevisiae могут включаться и выключаться медленно и независимо. Кроме того, мы раскрываем простую регуляторную стратегию, с помощью которой клетки могут контролировать долю клеток в различных состояниях экспрессии генов. Поскольку FLO11 кодирует белок клеточной стенки, ответственный за опосредование взаимодействий клетка-клетка и клетка-поверхность, этот контроль может буквально позволять естественным популяциям иметь контролируемую фракцию клеток «прилипать», в то время как другая фракция легко смывается.
cerevisiae могут включаться и выключаться медленно и независимо. Кроме того, мы раскрываем простую регуляторную стратегию, с помощью которой клетки могут контролировать долю клеток в различных состояниях экспрессии генов. Поскольку FLO11 кодирует белок клеточной стенки, ответственный за опосредование взаимодействий клетка-клетка и клетка-поверхность, этот контроль может буквально позволять естественным популяциям иметь контролируемую фракцию клеток «прилипать», в то время как другая фракция легко смывается.
Образец цитирования: Octavio LM, Gedeon K, Maheshri N (2009) Эпигенетическая и традиционная регуляция распределяется между активаторами FLO11 , позволяя настраивать гетерогенность на уровне популяции в ее выражении. PLoS Genet 5 (10): e1000673. https://doi.org/10.1371/journal.pgen.1000673
Редактор: Асифа Ахтар, Европейская лаборатория молекулярной биологии, Германия
Поступила: 17 апреля 2009 г . ; Одобрена: 2 сентября 2009 г .; Опубликовано: 2 октября 2009 г.
; Одобрена: 2 сентября 2009 г .; Опубликовано: 2 октября 2009 г.
Авторские права: © 2009 Octavio et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Эта работа была поддержана за счет средств стартапа MIT (Массачусетского технологического института) для NM, гранта NIH NIGMS на обучение NIH5R90DK071503 для LO и средств MIT UROP (Программа возможностей для исследований в бакалавриате) для KG. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Популяции микробных клеток используют ряд стратегий для быстрого создания фенотипического разнообразия в относительно короткие сроки [1], [2]. У некоторых микробов гены, известные как локусы непредвиденных обстоятельств, содержат тандемные повторы ДНК, рекомбинация которых приводит к включению или выключению экспрессии [3]. Другие генетические стратегии включают направленную рекомбинацию молчащих аллелей в конкретный активный локус, как в случае переключения типа спаривания у дрожжей и экспрессии поверхностного антигена у T. brucei [4], возбудитель африканской сонной болезни. Другой широко используемой стратегией, которая генерирует фенотипическую гетерогенность в популяциях клональных микробных клеток, является эпигенетическая регуляция генов. В отличие от генетических стратегий, это относится к наследуемому изменению экспрессии гена, которое не вызвано изменениями в последовательности основного гена. Например, паразит P. falciparum (малярия) и модельные организмы S. cerevisiae и E. coli используют эпигенетические механизмы для различной экспрессии антигенных белков клеточной поверхности [2] и, возможно, избегают иммунного надзора и / или выжить в непредсказуемо меняющейся среде.
У некоторых микробов гены, известные как локусы непредвиденных обстоятельств, содержат тандемные повторы ДНК, рекомбинация которых приводит к включению или выключению экспрессии [3]. Другие генетические стратегии включают направленную рекомбинацию молчащих аллелей в конкретный активный локус, как в случае переключения типа спаривания у дрожжей и экспрессии поверхностного антигена у T. brucei [4], возбудитель африканской сонной болезни. Другой широко используемой стратегией, которая генерирует фенотипическую гетерогенность в популяциях клональных микробных клеток, является эпигенетическая регуляция генов. В отличие от генетических стратегий, это относится к наследуемому изменению экспрессии гена, которое не вызвано изменениями в последовательности основного гена. Например, паразит P. falciparum (малярия) и модельные организмы S. cerevisiae и E. coli используют эпигенетические механизмы для различной экспрессии антигенных белков клеточной поверхности [2] и, возможно, избегают иммунного надзора и / или выжить в непредсказуемо меняющейся среде.
Многие эпигенетически регулируемые гены можно рассматривать как переключатели, поскольку они имеют два наследуемых состояния экспрессии: «ВКЛ» и «ВЫКЛ». Стабильный эпигенетический маркер поддерживает каждое состояние и может быть закодирован в цис или транс . Молекулярная основа локальных маркеров цис включает ковалентные модификации ДНК или ДНК-ассоциированных белков. К ним относятся метилирование ДНК [5] и модификации гистонов, которые определяют заглушенный гетерохроматин или активный эухроматин у эукариот [6].Глобальные маркеры транс и часто являются активностью фактора транскрипции; механизм стабильного, медленного переключения этих уровней — это положительные или двойные отрицательные петли обратной связи, которые генерируют наследуемые бистабильные состояния экспрессии генов, связанные с высокими или низкими уровнями активности факторов транскрипции [7] — [9]. Переключатели, использующие любую схему, реагируют на факторы окружающей среды, но неоднородность наблюдается даже при постоянных условиях окружающей среды, что свидетельствует о том, что переключатель может редко и случайным образом переключаться из-за колебаний во внутриклеточной среде. Эти две схемы также можно комбинировать. Например, в уропатогенном кишечнике E. coli экспрессия пилей, ассоциированных с пиелонефритом, регулируется эпигенетическим переключателем, который поддерживает свое состояние за счет как метилирования ДНК, так и петли положительной обратной связи [10].
Эти две схемы также можно комбинировать. Например, в уропатогенном кишечнике E. coli экспрессия пилей, ассоциированных с пиелонефритом, регулируется эпигенетическим переключателем, который поддерживает свое состояние за счет как метилирования ДНК, так и петли положительной обратной связи [10].
Контроль фенотипической гетерогенности, возможно, так же важен, как и ее быстрое образование. Гетерогенность, или шум, в традиционно регулируемой экспрессии генов хорошо изучена в последние годы. Исследования одиночных клеток и одиночных молекул показали, что активация генов происходит случайными, прерывистыми транскрипционными всплесками [11] — [14] из-за быстрых колебаний промотора (> одного раза за клеточный цикл) между неактивным (но компетентным) и активным состоянием промотора.С механистической точки зрения это чрезмерное упрощение, поскольку промотор, вероятно, принимает ряд различных состояний, включая связывание различных ген-специфических и общих транскрипционных механизмов, которые приводят к продуктивной транскрипции. Здесь активное состояние промотора можно рассматривать как состояние, при котором возможны быстрая инициация и повторная инициация. Напр., Для регулируемых промоторов, зависимых от Pol II РНК, инициация транскрипции часто ограничивает скорость и, следовательно, состояние активного промотора соответствует образованию преинициативного комплекса.Гетерогенность экспрессии, вызванная даже этими быстрыми колебаниями, может иметь последствия для фенотипа и приспособленности на уровне популяции [15].
Здесь активное состояние промотора можно рассматривать как состояние, при котором возможны быстрая инициация и повторная инициация. Напр., Для регулируемых промоторов, зависимых от Pol II РНК, инициация транскрипции часто ограничивает скорость и, следовательно, состояние активного промотора соответствует образованию преинициативного комплекса.Гетерогенность экспрессии, вызванная даже этими быстрыми колебаниями, может иметь последствия для фенотипа и приспособленности на уровне популяции [15].
Шум в экспрессии генов можно разделить в зависимости от того, является ли его источник внутренним или внешним по отношению к процессу экспрессии генов. Внутренний шум возникает из-за случайного характера химических превращений, включая события транскрипции и трансляции. Однако случайные всплески транскрипции, которые, как полагают, связаны с быстрыми флуктуациями промотора, происходящими в cis , по-видимому, являются доминирующим источником внутреннего шума у эукариот [16], [17]. Внешний шум возникает из-за межклеточной изменчивости транс- факторов, влияющих на экспрессию генов: например, общий и ген-специфический транскрипционный аппарат, количество рибосом и доступность тРНК или даже морфологию клетки. Два источника можно экспериментально различить с помощью двойного репортерного анализа, в котором две копии одного и того же промотора используются для управления различимыми вариантами флуоресцентного белка [18]. Внешний шум — это изменение уровня белка в разных клетках; Внутренний шум — это изменение уровня белка в одной и той же клетке.
Внешний шум возникает из-за межклеточной изменчивости транс- факторов, влияющих на экспрессию генов: например, общий и ген-специфический транскрипционный аппарат, количество рибосом и доступность тРНК или даже морфологию клетки. Два источника можно экспериментально различить с помощью двойного репортерного анализа, в котором две копии одного и того же промотора используются для управления различимыми вариантами флуоресцентного белка [18]. Внешний шум — это изменение уровня белка в разных клетках; Внутренний шум — это изменение уровня белка в одной и той же клетке.
То, как регуляторы контролируют кинетику собственных колебаний промотора, определяет результирующую гетерогенность экспрессии. Стохастические модели могут использоваться для прямой количественной оценки этой взаимосвязи [19]. Большинство регуляторов транскрипции, по-видимому, функционируют, модулируя частоту этих всплесков [19], [20], вероятно, в значительной степени за счет увеличения скорости инициации транскрипции. Следовательно, регуляторы не контролируют гетерогенность экспрессии независимо от уровня экспрессии. Фактически, гетерогенность находится под генетическим контролем, поскольку шумные промоторы имеют тенденцию иметь определенные характеристики: сильные TATA-боксы, высоко регулируемые и зависимые от активности ремоделирования хроматина [19] — [21].
Следовательно, регуляторы не контролируют гетерогенность экспрессии независимо от уровня экспрессии. Фактически, гетерогенность находится под генетическим контролем, поскольку шумные промоторы имеют тенденцию иметь определенные характеристики: сильные TATA-боксы, высоко регулируемые и зависимые от активности ремоделирования хроматина [19] — [21].
В то время как обычная регуляция генов включает быстрые колебания между неактивным (компетентным) и активным состояниями промотора, эпигенетическое подавление экспрессии генов включает медленные колебания (<один раз за клеточный цикл) между состоянием молчания и компетентным состоянием. Кинетика этих флуктуаций в переключателях, кодируемых trans , включающих петли обратной связи и связанных с бистабильной экспрессией генов, была подробно изучена [7], [22], [23]. И теория, и эксперимент предполагают, что внешние флуктуации фактора транс , которые преодолевают стабильность двух эпигенетических состояний, приводят к переключению [7]. Однако гораздо меньше известно о точной роли регуляторов в модуляции флуктуаций цис--закодированных переключателей, которые должны включать изменения в локальном состоянии промотора. Например, активаторы могут увеличивать усредненную по популяции экспрессию либо за счет стабилизации компетентного состояния, либо за счет дестабилизации состояния молчания. Результирующая неоднородность экспрессии диктуется особой кинетической ролью активатора.
Однако гораздо меньше известно о точной роли регуляторов в модуляции флуктуаций цис--закодированных переключателей, которые должны включать изменения в локальном состоянии промотора. Например, активаторы могут увеличивать усредненную по популяции экспрессию либо за счет стабилизации компетентного состояния, либо за счет дестабилизации состояния молчания. Результирующая неоднородность экспрессии диктуется особой кинетической ролью активатора.
В диплоидном организме эпигенетически регулируемый ген может демонстрировать четыре различных состояния экспрессии, если каждая копия переключается независимо.При глобальном кодировании обе копии реагируют на один и тот же глобальный фактор и должны переключаться коррелированным образом. Однако при локальном кодировании каждая копия может реагировать независимо, если колебание, вызывающее срабатывание переключателя, является молекулярным событием, которое происходит локально в одной копии. Фактически, недавнее исследование продемонстрировало случайное и независимое переключение двух копий репортерного гена, вставленного в локусы канонически замалчиваемого типа спаривания, HMR и HML в S. cerevisiae .Четыре различных состояния экспрессии наблюдались на фоне sir1 и , где SIR-белок-зависимое молчание этих локусов частично нарушено [24].
cerevisiae .Четыре различных состояния экспрессии наблюдались на фоне sir1 и , где SIR-белок-зависимое молчание этих локусов частично нарушено [24].
Множественные эпигенетические переключатели, закодированные цис , которые переключаются медленно и случайным образом, могут привести к комбинаторному взрыву состояний экспрессии и представляют собой мощную стратегию генерации фенотипического разнообразия. Используется ли в природе независимое переключение и как регулируются медленные колебания? S. cerevisiae Flo11p представляет собой белок адгезина клеточной стенки и член семейства генов FLO , важный в опосредовании межклеточных и гидрофобных взаимодействий с клеточной поверхностью [25].В дополнение к традиционной регуляции с помощью путей MAPK и PKA [26], [27] известно по крайней мере три механизма, генерирующих вариации адгезинов на клеточной поверхности: регуляция плоидности [28], частая рекомбинация тандемных повторов в генах адгезина [29]. ] и эпигенетическое молчание [30]. Молчание по FLO11 происходит независимо от SIR-белка и является как промотором, так и позиционно-специфичным [30]. Учитывая важность фенотипического разнообразия в адгезивном фенотипе и эпигенетического молчания на FLO11 , независимое переключение может представлять четвертый механизм для генерации вариации.
] и эпигенетическое молчание [30]. Молчание по FLO11 происходит независимо от SIR-белка и является как промотором, так и позиционно-специфичным [30]. Учитывая важность фенотипического разнообразия в адгезивном фенотипе и эпигенетического молчания на FLO11 , независимое переключение может представлять четвертый механизм для генерации вариации.
Промотор FLO11 размером 3,5 т.п.н. является одним из крупнейших в S. cerevisiae и регулируется многими факторами (Рис. 1), кинетическая роль которых неизвестна. Молчание FLO11 , как полагают, происходит посредством рекрутирования гистондеацетилазы Hda1p через репрессор Sfl1p посредством еще не определенного механизма [30]. Сайт связывания репрессора Sfl1p перекрывает сайт связывания активатора Flo8p [27]. Активация FLO11 через путь протеинкиназы A (PKA) приводит к фосфорилированию как Sfl1p, так и Flo8p.Хотя фосфорилирование отключает связывание Sfl1p, оно делает возможным связывание Flo8p [26], [27]. Дополнительные факторы транскрипции связываются непосредственно с этим промотором [26], [27], [31], включая регулируемые MAPK Ste12p / Tec1p и Phd1p. Эти три активатора нуждаются в Flo8p для активации и играют важную роль в определении общего уровня экспрессии [32]. Два активатора, Msn1p и Mss11p, не нуждаются в Flo8p для активации и действуют посредством плохо изученных механизмов, которые, по-видимому, не требуют связывания ДНК [33].Msn1p действует на больших расстояниях, дестабилизируя хроматин [34]; Mss11p имеет богатые глутамином активационные домены и может слабо связываться с Flo8p [32]. Все эти активаторы модулируют плазмидную экспрессию FLO11 , в контексте, где молчание не происходит [26], [27]. Однако их разнообразные биохимические роли могут указывать на разные кинетические и функциональные роли в эпигенетической регуляции FLO11 .
Дополнительные факторы транскрипции связываются непосредственно с этим промотором [26], [27], [31], включая регулируемые MAPK Ste12p / Tec1p и Phd1p. Эти три активатора нуждаются в Flo8p для активации и играют важную роль в определении общего уровня экспрессии [32]. Два активатора, Msn1p и Mss11p, не нуждаются в Flo8p для активации и действуют посредством плохо изученных механизмов, которые, по-видимому, не требуют связывания ДНК [33].Msn1p действует на больших расстояниях, дестабилизируя хроматин [34]; Mss11p имеет богатые глутамином активационные домены и может слабо связываться с Flo8p [32]. Все эти активаторы модулируют плазмидную экспрессию FLO11 , в контексте, где молчание не происходит [26], [27]. Однако их разнообразные биохимические роли могут указывать на разные кинетические и функциональные роли в эпигенетической регуляции FLO11 .
Рисунок 1. Сигналы от многих факторов транс сходятся в комплексном промоторе FLO11 .
Регуляторы транскрипции FLO11 . Позиции нуклеосом основаны на термодинамической модели занятости нуклеосом [37]. Сайты связывания являются приблизительными и основаны на литературе, но большинство сайтов не подтверждены напрямую. Также показаны три места, где была вставлена последовательность tetO. См. Основной текст, текст S1 и рисунок S1 для получения дополнительных сведений.
Позиции нуклеосом основаны на термодинамической модели занятости нуклеосом [37]. Сайты связывания являются приблизительными и основаны на литературе, но большинство сайтов не подтверждены напрямую. Также показаны три места, где была вставлена последовательность tetO. См. Основной текст, текст S1 и рисунок S1 для получения дополнительных сведений.
https://doi.org/10.1371/journal.pgen.1000673.g001
Здесь мы приводим доказательства того, что FLO11 действительно является эпигенетическим переключателем cis , и определяем кинетические роли трансфакторов в эпигенетическая и конвенциональная регуляция FLO11 .Внутри диплоидных дрожжей каждый локус переключается медленно, случайным образом и независимо, причем скорость переключения зависит от условий окружающей среды. Используя стохастическую кинетическую модель, мы делаем вывод о кинетической роли, которую разные регуляторы играют в медленных флуктуациях промотора, связанных с эпигенетическими переходами между молчащим и компетентным состоянием промотора, и быстрыми флуктуациями промотора, связанными с обычной активацией генов. Мы находим три класса регуляторов FLO11 : те, которые влияют на стабильность компетентного состояния, влияя на медленные колебания промотора; те, которые регулируют частоту всплесков транскрипции из-за быстрых колебаний промотора; и те, у которых есть обе функции.Более того, один синтетический активатор может имитировать каждый из этих трех классов в зависимости от расположения его сайта связывания ДНК. Поскольку кинетическая роль каждого регулятора определяет его влияние на неоднородность экспрессии, это может контролироваться выбором класса регулятора. Наконец, этанол контролирует степень молчания генов почти независимо от активации транскрипции посредством Flo8p, тем самым определяя, будет ли экспрессия FLO11 и отвечать ступенчатым или гетерогенным образом на другие сигналы.
Мы находим три класса регуляторов FLO11 : те, которые влияют на стабильность компетентного состояния, влияя на медленные колебания промотора; те, которые регулируют частоту всплесков транскрипции из-за быстрых колебаний промотора; и те, у которых есть обе функции.Более того, один синтетический активатор может имитировать каждый из этих трех классов в зависимости от расположения его сайта связывания ДНК. Поскольку кинетическая роль каждого регулятора определяет его влияние на неоднородность экспрессии, это может контролироваться выбором класса регулятора. Наконец, этанол контролирует степень молчания генов почти независимо от активации транскрипции посредством Flo8p, тем самым определяя, будет ли экспрессия FLO11 и отвечать ступенчатым или гетерогенным образом на другие сигналы.
Результаты
FLO11 переключается между состояниями без звука и компетентностью независимо на каждом локусе В условиях плохого питания FLO11 частично замалчивается в гаплоидных клетках и неоднороден по экспрессии. Члены этой популяции способны обратимо переходить между выключенным (без звука) и включенным (компетентным) состоянием [30]. Чтобы определить, связаны ли переходы с флуктуацией цис или транс , мы использовали двойной репортерный анализ, заменив две копии ORF FLO11 в диплоидных дрожжах на отдельный вариант флуоресцентного белка (Venus YFP и Cerulean CFP) (рисунок 2A).Важно отметить, что мы проверили независимость и эквивалентность двух репортеров в отношении присутствия другого репортера (Рисунок S2 и Рисунок S3). При выращивании в среде с плохими источниками углерода, включая этанол, глицерин, галактозу и раффинозу, мы наблюдали все четыре возможных состояния экспрессии (рисунок 2B, данные не показаны). Поскольку эндогенный Flo11p не присутствует в штамме с двумя репортерами, мы подтвердили, что Flo11p не влияет на экспрессию на промоторе FLO11 двумя способами.Сначала мы добавили плазмиду, конститутивно экспрессирующую FLO11 , и не обнаружили значительного влияния на экспрессию флуоресцентного белка (данные не показаны). Во-вторых, мы сравнили экспрессию флуоресцентного белка в штамме с двумя репортерами со штаммами, в которых только один аллель FLO11 был заменен флуоресцентным белком. Не было разницы в уровнях экспрессии (рисунок S2 и рисунок S3).
Во-вторых, мы сравнили экспрессию флуоресцентного белка в штамме с двумя репортерами со штаммами, в которых только один аллель FLO11 был заменен флуоресцентным белком. Не было разницы в уровнях экспрессии (рисунок S2 и рисунок S3).
Рисунок 2. Смешанные состояния экспрессии и независимое переключение в локусе FLO11 .
(A) Двойной репортерный анализ. Каждый аллель FLO11 включается и выключается медленно с одинаковой скоростью λ и γ , потому что два репортера эквивалентны. (B) Смешанные состояния выражения. Штамм с двойным репортером, выращенный в богатой среде (без глюкозы) с добавлением 1% этанола и 2% глицерина (наложение ложного цвета CFP = красный, YFP = зеленый). Видны все четыре возможных состояния выражения. (C) Скорость перехода. Эквивалентность репортеров подразумевает λ 1 = λ 2 , λ 3 = λ 4 , γ 1 = γ 2 , γ 3 = γ 4 . Независимое переключение подразумевает λ 1 = λ 3 и γ 1 = γ 3 . (D) Скорость перехода из ВЫКЛ в ВКЛ различных состояний экспрессии: λ 1 (X), λ 2 (X), λ 3 (•), λ 4 ( •). Каждый маркер представляет долю клеток, в которых наблюдается переключение в конкретный момент времени, а розовая кривая такая же, как и аппроксимирующая кривая на рисунке 3А.(E) То же, что и в (D), но для скоростей перехода из ВКЛ в ВЫКЛ: γ 1 (X), γ 2 (X), γ 3 (•), γ 4 (•). Синяя кривая такая же, как аппроксимирующая кривая на рисунке 3A. (D) и (E) демонстрируют, что скорость перехода на одном аллеле не зависит от состояния другого аллеля. Даже нулевая гипотеза о том, что γ 2 и γ 4 эквивалентны, не может быть отклонена на уровне значимости 5% (двусторонний T-критерий, p = 0.
Независимое переключение подразумевает λ 1 = λ 3 и γ 1 = γ 3 . (D) Скорость перехода из ВЫКЛ в ВКЛ различных состояний экспрессии: λ 1 (X), λ 2 (X), λ 3 (•), λ 4 ( •). Каждый маркер представляет долю клеток, в которых наблюдается переключение в конкретный момент времени, а розовая кривая такая же, как и аппроксимирующая кривая на рисунке 3А.(E) То же, что и в (D), но для скоростей перехода из ВКЛ в ВЫКЛ: γ 1 (X), γ 2 (X), γ 3 (•), γ 4 (•). Синяя кривая такая же, как аппроксимирующая кривая на рисунке 3A. (D) и (E) демонстрируют, что скорость перехода на одном аллеле не зависит от состояния другого аллеля. Даже нулевая гипотеза о том, что γ 2 и γ 4 эквивалентны, не может быть отклонена на уровне значимости 5% (двусторонний T-критерий, p = 0. 28), равно как и нулевая гипотеза об идентичности их распределений (двусторонний тест KS, p = 0,47).
28), равно как и нулевая гипотеза об идентичности их распределений (двусторонний тест KS, p = 0,47).
https://doi.org/10.1371/journal.pgen.1000673.g002
Если каждый аллель переключается независимо, то в устойчивом состоянии доля клеток в каждом состоянии экспрессии определяется как p 2 (оба включены ), (1− p ) 2 (оба выключены) или 2 p (1− p ) (смешанные ВКЛ / ВЫКЛ и ВЫКЛ / ВКЛ), где p — это доля ячеек с конкретный аллель ВКЛ.Обратите внимание, что p идентичен как для экспрессии YFP, так и для CFP, потому что аллели эквивалентны. Мы смогли убедиться, что профиль экспрессии популяции достиг стабильного состояния (Рисунок 3A и Рисунок S4). Однако наивная классификация состояния экспрессии, основанная на сравнении уровня флуоресценции клетки с фоном, неверна, поскольку она не учитывает длительное время жизни флуоресцентных белков, которое скрывает истинное состояние экспрессии промотора. Таким образом, мы непосредственно измерили восемь скоростей перехода путем мониторинга в реальном времени экспрессии FLO11 в отдельных клетках, выращенных в микрофлюидной камере при постоянных условиях (подробности в тексте S1).Было обнаружено, что все четыре скорости перехода из ON в OFF и из OFF в ON (рис. 2C – 2E) неразличимы, демонстрируя, что каждый аллель переключался независимо. Кроме того, доля включенных или выключенных ячеек хорошо согласуется с одной экспонентой, подтверждая, что каждый переход надлежащим образом отнесен к реакции псевдопервого порядка. Переключение не коррелировало со стадией клеточного цикла (рисунок S7).
Таким образом, мы непосредственно измерили восемь скоростей перехода путем мониторинга в реальном времени экспрессии FLO11 в отдельных клетках, выращенных в микрофлюидной камере при постоянных условиях (подробности в тексте S1).Было обнаружено, что все четыре скорости перехода из ON в OFF и из OFF в ON (рис. 2C – 2E) неразличимы, демонстрируя, что каждый аллель переключался независимо. Кроме того, доля включенных или выключенных ячеек хорошо согласуется с одной экспонентой, подтверждая, что каждый переход надлежащим образом отнесен к реакции псевдопервого порядка. Переключение не коррелировало со стадией клеточного цикла (рисунок S7).
Рис. 3. Метод вывода кинетики переключения из статического установившегося распределения.
(A) (слева) Эволюция во времени популяционного распределения экспрессии YFP из штамма с двумя репортерами, растущего в YP 1% этанола, 2% глицерина в микрофлюидной камере в течение 20 часов. Цветная полоса указывает долю ячеек ( n = 230 с течением времени). Перед переносом в микрофлюидную камеру этот штамм рос в идентичных условиях в жидкой культуре. Распределение меняется на ранней стадии из-за небольшого исходного размера выборки ( n = 10). (Справа) Граничные скорости перехода между состояниями ВКЛ и ВЫКЛ.Синие / розовые точки указывают на долю клеток, включенных / выключенных при рождении и наблюдаемых для выключения / включения. Соответствующие кривые соответствуют модели экспоненциально распределенного времени переключения с ВКЛ на ВЫКЛ и ВЫКЛ на ВКЛ, с скорректированными скоростями, показанными рядом с графиком. Планки погрешностей соответствуют 3 s.d. от среднего, рассчитанного с помощью бутстрап-анализа. Аналогичные результаты получаются при сосредоточении внимания на экспрессии CFP (см. Рисунок S5 и рисунок S6). (B) Модель с тремя состояниями активации FLO11 , показывающая разделение шкал времени между эпигенетической (молчание) и традиционной регуляцией.Когда присутствуют медленные переходы, связанные с молчанием, быстрые переходы транскрипционного взрыва могут быть объединены в одну скорость μ.
Перед переносом в микрофлюидную камеру этот штамм рос в идентичных условиях в жидкой культуре. Распределение меняется на ранней стадии из-за небольшого исходного размера выборки ( n = 10). (Справа) Граничные скорости перехода между состояниями ВКЛ и ВЫКЛ.Синие / розовые точки указывают на долю клеток, включенных / выключенных при рождении и наблюдаемых для выключения / включения. Соответствующие кривые соответствуют модели экспоненциально распределенного времени переключения с ВКЛ на ВЫКЛ и ВЫКЛ на ВКЛ, с скорректированными скоростями, показанными рядом с графиком. Планки погрешностей соответствуют 3 s.d. от среднего, рассчитанного с помощью бутстрап-анализа. Аналогичные результаты получаются при сосредоточении внимания на экспрессии CFP (см. Рисунок S5 и рисунок S6). (B) Модель с тремя состояниями активации FLO11 , показывающая разделение шкал времени между эпигенетической (молчание) и традиционной регуляцией.Когда присутствуют медленные переходы, связанные с молчанием, быстрые переходы транскрипционного взрыва могут быть объединены в одну скорость μ.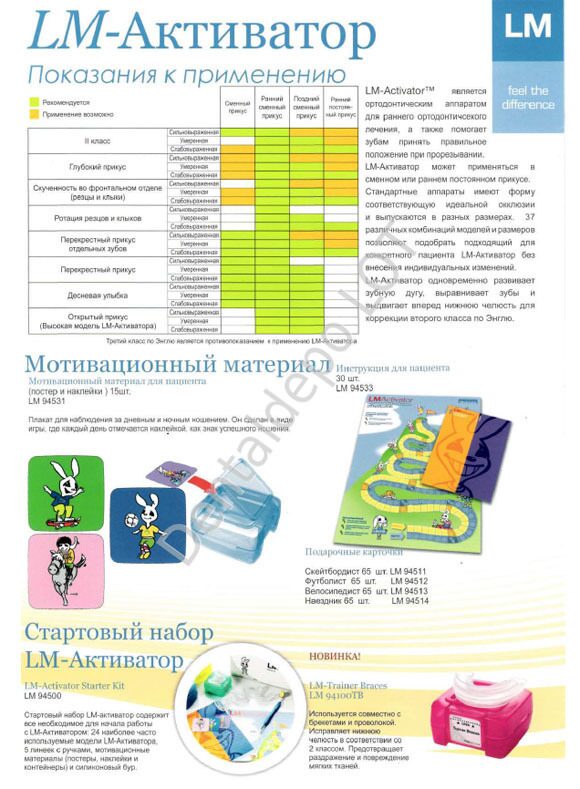 Затем модель сворачивается в модель с двумя состояниями в (C). (C) (Слева) Статическое распределение флуоресценции YFP штамма с двумя репортерами, выращенного в тех же условиях среды, что и A, но в планшетах с глубокими лунками, а не в микрофлюидном устройстве. Частота переходов, полученная из этого снимка с использованием стохастической кинетической модели (справа), близко согласуется с данными, полученными с помощью покадровой микроскопии.(D) Модуляция скорости переключения. Прогноз стохастической кинетической модели доли клеток в состоянии смешанной экспрессии соответствует независимому переключению (заданному как 2 p (1-p) , что соответствует серой линии) для ряда условий. Планки погрешностей (ось x) взяты из 95% доверительных интервалов от MLE-соответствия частоты переключения для оценки истинной доли включенных ячеек; планки погрешностей (ось Y) связаны с ошибками в оценке порогового значения флуоресценции для автофлуоресценции (см. текст S1).
Затем модель сворачивается в модель с двумя состояниями в (C). (C) (Слева) Статическое распределение флуоресценции YFP штамма с двумя репортерами, выращенного в тех же условиях среды, что и A, но в планшетах с глубокими лунками, а не в микрофлюидном устройстве. Частота переходов, полученная из этого снимка с использованием стохастической кинетической модели (справа), близко согласуется с данными, полученными с помощью покадровой микроскопии.(D) Модуляция скорости переключения. Прогноз стохастической кинетической модели доли клеток в состоянии смешанной экспрессии соответствует независимому переключению (заданному как 2 p (1-p) , что соответствует серой линии) для ряда условий. Планки погрешностей (ось x) взяты из 95% доверительных интервалов от MLE-соответствия частоты переключения для оценки истинной доли включенных ячеек; планки погрешностей (ось Y) связаны с ошибками в оценке порогового значения флуоресценции для автофлуоресценции (см. текст S1).
https://doi. org/10.1371/journal.pgen.1000673.g003
org/10.1371/journal.pgen.1000673.g003
Используя стохастическую кинетическую модель, статические распределения могут раскрыть кинетическую информацию
Покадровая микроскопия обеспечивает точное определение медленных скоростей эпигенетических переходов и доли каждого состояния экспрессии, но это экспериментально сложная задача и низкая пропускная способность. Таким образом, после определения того, что скорости перехода были точно описаны как первого порядка, мы разработали способ вывести эти скорости непосредственно из статических снимков, учитывая длительный срок жизни флуоресцентных репортеров.Модели с двумя состояниями широко используются для моделирования более быстрых колебаний промотора, связанных с обычной регуляцией генов [16], [21], [35]. В таких моделях промотор может переходить из неактивного, но компетентного состояния в активное состояние, которое приводит к транскрипции. Многие эукариотические гены, по-видимому, находятся в компетентном состоянии с редкими переходами в короткоживущее активное состояние, что приводит к «всплеску» транскрипции.
Наблюдаемые вариации промотора FLO11 можно разделить на внутренний и внешний компоненты.Промотор FLO11 подвержен как быстрым внутренним колебаниям, так и медленному эпигенетическому переходу, как показано в расширенной модели с тремя состояниями на рисунке 3B. Внешний шум также способствует изменению уровней экспрессии FLO11 от клетки к клетке, когда промотор не заглушен. Однако, когда промотор (частично) заглушен, преобладающий источник вариаций в FLO11 или репортерной экспрессии возникает из медленного эпигенетического перехода между заглушенным и компетентным состояниями из-за (1) меньшей величины быстрых внутренних и внешних флуктуаций и тот факт, что (2) более быстрые флуктуации (<1 поколение клеток) более полно усредняются по времени долгоживущим репортером по сравнению с медленным переходом (> 1 поколение клеток).Следовательно, мы можем объединить быстрые скорости перехода ( λ ‘, γ ‘, μ ′) в общую скорость транскрипции μ и игнорировать внешние флуктуации. Экспрессия гена теперь может быть описана с использованием обычно используемой модели с двумя состояниями: где x i s — количество репортерного белка, μ — (сосредоточенная) скорость продукции белка, δ — скорость деградации белка. (здесь скорость роста клеток), а f (t) — это «случайный телеграфный процесс», который принимает значения 0 или 1, соответствующие отключенному или активному состоянию промотора, с экспоненциально распределенным временем между событиями переключения (рис. 3C).Это стохастическое уравнение было решено аналитически, чтобы получить бета-распределение для белка с номером x в стационарном состоянии [35]. Медленные скорости эпигенетических переходов, λ и γ , соответствуют тем, которые были измерены в эксперименте по времени. Чтобы сделать вывод об этих скоростях, мы предполагаем, что наше измеренное распределение белка x является устойчивым (рисунок S4), и подгоняем его к бета-распределению, используя значение μ , основанное на уровне экспрессии популяции ON в бимодальном состоянии (параметр δ , скорость роста клеток, измеряется напрямую — см.
Экспрессия гена теперь может быть описана с использованием обычно используемой модели с двумя состояниями: где x i s — количество репортерного белка, μ — (сосредоточенная) скорость продукции белка, δ — скорость деградации белка. (здесь скорость роста клеток), а f (t) — это «случайный телеграфный процесс», который принимает значения 0 или 1, соответствующие отключенному или активному состоянию промотора, с экспоненциально распределенным временем между событиями переключения (рис. 3C).Это стохастическое уравнение было решено аналитически, чтобы получить бета-распределение для белка с номером x в стационарном состоянии [35]. Медленные скорости эпигенетических переходов, λ и γ , соответствуют тем, которые были измерены в эксперименте по времени. Чтобы сделать вывод об этих скоростях, мы предполагаем, что наше измеренное распределение белка x является устойчивым (рисунок S4), и подгоняем его к бета-распределению, используя значение μ , основанное на уровне экспрессии популяции ON в бимодальном состоянии (параметр δ , скорость роста клеток, измеряется напрямую — см. Текст S1).
Текст S1).
Мы протестировали этот метод двумя разными способами. Во-первых, мы использовали стационарное распределение белков в экспериментах с интервальной съемкой для оценки скорости перехода и обнаружили тесное соответствие между предполагаемыми скоростями и теми, которые непосредственно измерялись в промежутке времени (рис. 3A и 3C). Во-вторых, эта модель позволяет правильно оценить долю клеток, которые появляются в статическом распределении, но фактически выключены из-за длительного срока службы флуоресцентного репортера (подробности в тексте S1). Мы применили эту поправку к статическим снимкам клеток, выращенных в различных условиях.Хотя доля клеток в каждом состоянии экспрессии варьировалась, общая статистика всегда соответствовала независимому переключению на каждом промоторе (рис. 3D). Следовательно, вышестоящая сигнальная сеть может отображать входные данные окружающей среды в конкретную смесь состояний экспрессии посредством модуляции скорости перехода.
Стратегия определения того, как регулирующие органы влияют на скорость перехода
В конечном счете, сигналы окружающей среды модулируют эпигенетическую регуляцию FLO11 через нижестоящие регуляторы. Эффект этих регуляторов как на средний уровень экспрессии, так и на гетерогенность экспрессии кратко и количественно описывается их влиянием на скорость перехода в модели с тремя состояниями (Рисунок 3B). Поэтому мы решили титровать транс-факторов и измерить количественный ответ промотора FLO11 на уровне отдельных клеток в сотнях клеток с помощью флуоресцентной микроскопии с использованием анализа с двумя репортерами. Для каждого состояния и штамма мы всегда выращивали клетки для> 10 удвоений, последовательно разбавляя их по мере необходимости, чтобы поддерживать низкую плотность и обеспечить достижение стабильного состояния (более подробная информация в тексте S1).
Эффект этих регуляторов как на средний уровень экспрессии, так и на гетерогенность экспрессии кратко и количественно описывается их влиянием на скорость перехода в модели с тремя состояниями (Рисунок 3B). Поэтому мы решили титровать транс-факторов и измерить количественный ответ промотора FLO11 на уровне отдельных клеток в сотнях клеток с помощью флуоресцентной микроскопии с использованием анализа с двумя репортерами. Для каждого состояния и штамма мы всегда выращивали клетки для> 10 удвоений, последовательно разбавляя их по мере необходимости, чтобы поддерживать низкую плотность и обеспечить достижение стабильного состояния (более подробная информация в тексте S1).
Чтобы получить скорости перехода, мы аппроксимируем измеренные распределения флуоресценции, возникающие при каждом титровании, с бета-распределением, решением упрощенной модели с двумя состояниями. Как описано ранее, модель с двумя состояниями, которая объединяет быстрые переходы, строго применима только тогда, когда доминируют медленные эпигенетические переходы, связанные с молчанием. Четырехквадрантный график на рисунке 4A суммирует качественный отклик на уровне популяции, заданный бета-распределением для различных комбинаций λ и γ .Каждый квадрант соответствует режимам, в которых λ и γ медленнее или быстрее, чем скорость деления ( δ ). Эпигенетическая регуляция происходит по определению в нижнем левом квадранте, когда и λ , и γ медленнее, чем скорость деления ( λ∼δ и γ∼δ <1). Экспрессия может полностью выключиться, если активное состояние дестабилизировано ( γ увеличивается, сдвиг в нижний правый квадрант) или состояние молчания стабилизируется ( λ уменьшается, бимодальная экспрессия с исчезающе меньшим процентом клеток ВКЛЮЧЕНА).Противоположные изменения в λ и γ полностью включают выражение (и может привести к смещению в верхний правый квадрант).
Четырехквадрантный график на рисунке 4A суммирует качественный отклик на уровне популяции, заданный бета-распределением для различных комбинаций λ и γ .Каждый квадрант соответствует режимам, в которых λ и γ медленнее или быстрее, чем скорость деления ( δ ). Эпигенетическая регуляция происходит по определению в нижнем левом квадранте, когда и λ , и γ медленнее, чем скорость деления ( λ∼δ и γ∼δ <1). Экспрессия может полностью выключиться, если активное состояние дестабилизировано ( γ увеличивается, сдвиг в нижний правый квадрант) или состояние молчания стабилизируется ( λ уменьшается, бимодальная экспрессия с исчезающе меньшим процентом клеток ВКЛЮЧЕНА).Противоположные изменения в λ и γ полностью включают выражение (и может привести к смещению в верхний правый квадрант).
Рис. 4. Три различных кинетических роли регуляторов FLO11 .
(A) Качественная форма бета-распределения для различных значений скоростей перехода ВЫКЛ → ВКЛ ( λ / δ ) и ВКЛ → ВЫКЛ ( γ / δ ) (нормированные относительно роста скорость δ ). Когда обе скорости ниже, чем рост (нижний левый квадрант), они характеризуют медленные эпигенетические переходы между безмолвным и компетентным состояниями.Распределение экспрессии является бимодальным, представляя стабильные популяции ON и OFF. Эти показатели могут быть определены путем измерения распределения экспрессии с помощью флуоресцентной микроскопии и подгонки к бета-распределению. Для унимодальных распределений ON эпигенетическое молчание больше не происходит. Если бы присутствовали только быстрые внутренние флуктуации между компетентным и активным состояниями промотора, была бы применима та же модель с двумя состояниями, но теперь можно прогнозировать быстрые скорости переходов и унимодальные распределения (верхняя половина графика).Однако, поскольку внешние флуктуации также имеют значение, прямая подгонка измеренных унимодальных распределений не дает , а не , чтобы получить быстрые скорости перехода (см. Основной текст и текст S1 для подробностей). (B) Типичные гистограммы флуоресценции трех классов активаторов. (Вверху) Tec1p, титрованный на фоне дикого типа в SD ura-; Flo8p титруют в фоновом растворе flo8Δ в SD ура- + 1% этаноле; Msn1p титруют на фоне дикого типа в SD ura-. (Внизу) rtTA, титрованная в штамме с tetO при -350 (сайт, закрытый нуклеосомами), при -1160 (сайт перекрывает сайт связывания Sfl1p) и при -1470 (сайт, расположенный непосредственно перед областью, свободной от нуклеосом -1200).Гистограммы получены с помощью флуоресцентной микроскопии (количество клеток> 300). Распределение флуоресценции штамма OFF (Y92), используемого для измерения аутофлуоресценции, показано вверху каждого графика. (C) Кинетические роли регуляторов. Повышение уровней различных активаторов FLO11 снижает γ , стабилизируя активное состояние без значительного изменения λ . Активаторы класса I не могут значительно уменьшить γ (синий). Активаторы класса II могут сдвигать скорость перехода в нижний левый квадрант, который соответствует частично подавленной бимодальной экспрессии (розовый).
(Вверху) Tec1p, титрованный на фоне дикого типа в SD ura-; Flo8p титруют в фоновом растворе flo8Δ в SD ура- + 1% этаноле; Msn1p титруют на фоне дикого типа в SD ura-. (Внизу) rtTA, титрованная в штамме с tetO при -350 (сайт, закрытый нуклеосомами), при -1160 (сайт перекрывает сайт связывания Sfl1p) и при -1470 (сайт, расположенный непосредственно перед областью, свободной от нуклеосом -1200).Гистограммы получены с помощью флуоресцентной микроскопии (количество клеток> 300). Распределение флуоресценции штамма OFF (Y92), используемого для измерения аутофлуоресценции, показано вверху каждого графика. (C) Кинетические роли регуляторов. Повышение уровней различных активаторов FLO11 снижает γ , стабилизируя активное состояние без значительного изменения λ . Активаторы класса I не могут значительно уменьшить γ (синий). Активаторы класса II могут сдвигать скорость перехода в нижний левый квадрант, который соответствует частично подавленной бимодальной экспрессии (розовый). Flo8p имеет менее стабильное состояние глушителя по сравнению с активаторами класса II. Похоже, что при критическом значении γ регуляторы отменяют молчание, и ответ попадает в верхний левый квадрант. (D) Титрование синтетического активатора. Титрование синтетических активаторов имитирует три класса активаторов в зависимости от местоположения сайта связывания. Все титрования (B, C и D) проводились в SD ura- за исключением Sfl1p и Flo8p, где был добавлен 1% этанол. Планки погрешностей представляют собой 95% доверительные интервалы (для соответствия экспериментальных данных бета-распределению для одного значения μ — подробности в тексте S1).
Flo8p имеет менее стабильное состояние глушителя по сравнению с активаторами класса II. Похоже, что при критическом значении γ регуляторы отменяют молчание, и ответ попадает в верхний левый квадрант. (D) Титрование синтетического активатора. Титрование синтетических активаторов имитирует три класса активаторов в зависимости от местоположения сайта связывания. Все титрования (B, C и D) проводились в SD ura- за исключением Sfl1p и Flo8p, где был добавлен 1% этанол. Планки погрешностей представляют собой 95% доверительные интервалы (для соответствия экспериментальных данных бета-распределению для одного значения μ — подробности в тексте S1).
https://doi.org/10.1371/journal.pgen.1000673.g004
Если эпигенетическая регуляция потеряна, уровни экспрессии все еще могут изменяться из-за более быстрых колебаний промотора. Модель с двумя состояниями точно описывает внутренние (но не внешние) флуктуации, вызванные переходами между компетентным и активным состояниями промотора.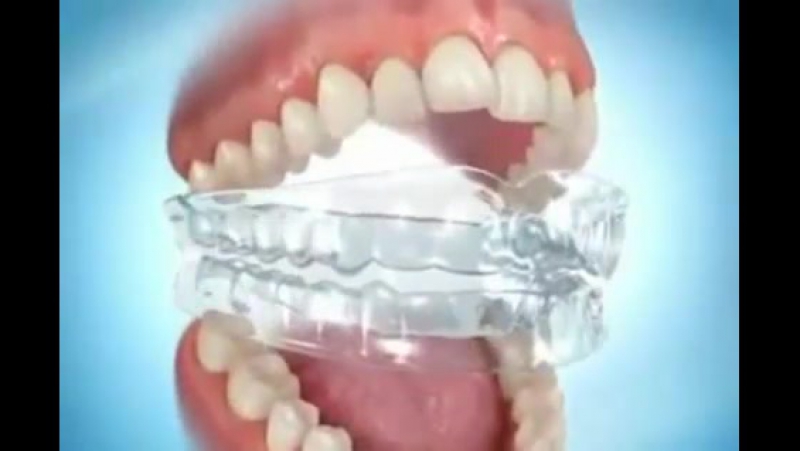 Например, обычно регулируемый (но репрессированный) ген может быть выключен и находиться в нижнем правом квадранте. Активация приводит к увеличению частоты всплесков (увеличивается на λ ‘) и постепенному унимодальному распределению в верхнем правом квадранте.Важно отметить, что невозможно различить обычную репрессию и эпигенетическое молчание в любой популяции в нижнем правом квадранте, где экспрессия полностью отключена. Более того, подгонка распределений экспрессии флуоресценции, генерируемых одним управляемым промотором FLO11 репортером, который обычно регулируется, не дает быстрых скоростей перехода промотора λ ‘ и γ’ , потому что здесь внешние колебания значительны. Внешний шум возникает из-за межклеточной изменчивости таких факторов, как морфология, количество рибосом и / или вышестоящие компоненты в регуляторном пути FLO11 , и влияет на оба промотора в одной и той же клетке коррелированным образом.Чтобы правильно измерить быстрые скорости перехода, связанные с традиционным регулированием, необходимо проанализировать собственный шум, чтобы определить частоту пакета ( λ ′ ) и размер пакета ( μ ′ / γ ′ ) (см.
Например, обычно регулируемый (но репрессированный) ген может быть выключен и находиться в нижнем правом квадранте. Активация приводит к увеличению частоты всплесков (увеличивается на λ ‘) и постепенному унимодальному распределению в верхнем правом квадранте.Важно отметить, что невозможно различить обычную репрессию и эпигенетическое молчание в любой популяции в нижнем правом квадранте, где экспрессия полностью отключена. Более того, подгонка распределений экспрессии флуоресценции, генерируемых одним управляемым промотором FLO11 репортером, который обычно регулируется, не дает быстрых скоростей перехода промотора λ ‘ и γ’ , потому что здесь внешние колебания значительны. Внешний шум возникает из-за межклеточной изменчивости таких факторов, как морфология, количество рибосом и / или вышестоящие компоненты в регуляторном пути FLO11 , и влияет на оба промотора в одной и той же клетке коррелированным образом.Чтобы правильно измерить быстрые скорости перехода, связанные с традиционным регулированием, необходимо проанализировать собственный шум, чтобы определить частоту пакета ( λ ′ ) и размер пакета ( μ ′ / γ ′ ) (см. Текст S1).
Текст S1).
Три класса регуляторов экспрессии FLO11
Чтобы отделить комплекс вышестоящих сигнальных событий, происходящих на промоторе (Рис. 1), индивидуальные факторы trans были гетерологично экспрессированы под контролем индуцируемого доксициклином промотора [36].Поскольку Sfl1p и Flo8p регулируются посттрансляционно, нам нужен был способ настроить их относительную силу. Мы выбрали этанол, поскольку добавление этанола активирует экспрессию FLO11 Flo8p-зависимым образом (см. Ниже). Все титрования фактора транс проводили в SD ura- или SD ura- с этанолом. Титрование Sfl1p, Flo8p и Mss11p проводили на фоне sfl1Δ , flo8Δ или mss11Δ соответственно.
Для каждой точки титрования мы вывели скорости перехода λ и γ , используя модель с двумя состояниями.Чтобы суммировать влияние различных активаторов и Sfl1p на стабильность безмолвных и компетентных состояний, мы наносим ряд значений ( γ , λ ), определенных на четырех квадрантном графике на рисунке 4A. В SD ura-, FLO11 выключен, что соответствует правому нижнему квадранту. Основываясь на ответе промотора FLO11 (рис. 4B и 4C), мы сгруппировали активаторы в 3 класса. Добавление трех активаторов класса I, Tec1p, Ste12p и Phd1p, по-видимому, слабо стабилизирует компетентное состояние, но экспрессия остается чрезвычайно низкой.Активаторы класса II, Msn1p и Mss11p, стабилизировали компетентное состояние, уменьшая γ и входя в гетерогенную область, где преобладают медленные флуктуации промотора. При некотором критическом значении γ состояние молчания быстро дестабилизируется, и вся популяция включается. Flo8p представляет собой особый класс и был титрован в условиях этанола, где предположительно некоторая фракция Flo8p теперь фосфорилирована и активна. Популяционный ответ занимает промежуточное положение между активаторами класса I и класса II, но ближе к классу II.Sfl1p оказывает прямо противоположный эффект в тех же условиях этанола, что согласуется с антагонистической ролью Sfl1p и Flo8p через их перекрывающиеся сайты связывания и как отрицательно и положительно регулируются PKA, соответственно.
В SD ura-, FLO11 выключен, что соответствует правому нижнему квадранту. Основываясь на ответе промотора FLO11 (рис. 4B и 4C), мы сгруппировали активаторы в 3 класса. Добавление трех активаторов класса I, Tec1p, Ste12p и Phd1p, по-видимому, слабо стабилизирует компетентное состояние, но экспрессия остается чрезвычайно низкой.Активаторы класса II, Msn1p и Mss11p, стабилизировали компетентное состояние, уменьшая γ и входя в гетерогенную область, где преобладают медленные флуктуации промотора. При некотором критическом значении γ состояние молчания быстро дестабилизируется, и вся популяция включается. Flo8p представляет собой особый класс и был титрован в условиях этанола, где предположительно некоторая фракция Flo8p теперь фосфорилирована и активна. Популяционный ответ занимает промежуточное положение между активаторами класса I и класса II, но ближе к классу II.Sfl1p оказывает прямо противоположный эффект в тех же условиях этанола, что согласуется с антагонистической ролью Sfl1p и Flo8p через их перекрывающиеся сайты связывания и как отрицательно и положительно регулируются PKA, соответственно..JPG)
Чтобы определить, коррелировал ли класс активатора с положением или доступностью сайта связывания, мы применили модель занятости нуклеосом in silico [37] и выполнили картирование микрококковой нуклеазы (рисунок 1 и рисунок S8) промотора FLO11 .Оба метода показали, что область -1200, содержащая перекрывающиеся сайты связывания для Sfl1p и Flo8p, не содержит нуклеосом. В отличие от этого, сайты связывания активаторов класса I находятся в закрытых нуклеосомами областях. Активаторы класса II, как известно, не связывают ДНК, но являются мощными активаторами, при этом Msn1p обладает известной способностью рекрутировать механизмы ремоделирования хроматина [34].
Синтетический активатор имитирует каждый класс активаторов в зависимости от положения его сайта связывания
Достаточно ли положения, доступности и / или конкуренции с Sfl1p для определения класса активатора? Если это так, синтетический активатор может иметь качественно разные регуляторные профили в зависимости от положения сайта связывания.![]() Мы сконструировали штаммы дрожжей с двумя репортерами с последовательностью tetO длиной 19 п.н., вставленной в 3 разных места в промоторе FLO11 . Первое местоположение было на -350 в области, закрытой нуклеосомами, рядом с ТАТА-боксом и сайтом начала транскрипции. Второе местоположение было на -1470, на внешнем крае нуклеосомы перед сайтом связывания Sfl1p и далеко от корового промотора. Третье местоположение было на -1160, в свободной от нуклеосом области, непосредственно перекрывающей сайт связывания Sfl1p.Sfl1p связывается как димер на двух сайтах [27], [38], поэтому мы заменили 19 п.н. промоторной последовательности между двумя сайтами на последовательность tetO, чтобы сохранить расстояние, охватываемое двумя сайтами. TetO связывается с rtTA, синтетическим активатором, который содержит сильный кислотный активационный домен VP16 [36], который, как известно, рекрутирует комплекс SAGA в дрожжах [39]. Мы титровали rtTA в этих трех штаммах, выращенных в SD ura- (все изначально выключены).
Мы сконструировали штаммы дрожжей с двумя репортерами с последовательностью tetO длиной 19 п.н., вставленной в 3 разных места в промоторе FLO11 . Первое местоположение было на -350 в области, закрытой нуклеосомами, рядом с ТАТА-боксом и сайтом начала транскрипции. Второе местоположение было на -1470, на внешнем крае нуклеосомы перед сайтом связывания Sfl1p и далеко от корового промотора. Третье местоположение было на -1160, в свободной от нуклеосом области, непосредственно перекрывающей сайт связывания Sfl1p.Sfl1p связывается как димер на двух сайтах [27], [38], поэтому мы заменили 19 п.н. промоторной последовательности между двумя сайтами на последовательность tetO, чтобы сохранить расстояние, охватываемое двумя сайтами. TetO связывается с rtTA, синтетическим активатором, который содержит сильный кислотный активационный домен VP16 [36], который, как известно, рекрутирует комплекс SAGA в дрожжах [39]. Мы титровали rtTA в этих трех штаммах, выращенных в SD ura- (все изначально выключены). Каждое местоположение функционально имитирует реакцию соответствующего класса активаторов (рис. 4B и 4D).Важно отметить, что стабильность состояния молчания снижается для третьего положения tetO по сравнению со вторым местом tetO. Это происходило даже в отсутствие этанола, что позволяет предположить, что разница в стабильности в молчаливом состоянии между активаторами класса II и Flo8p не является следствием альтернативного специфического для этанола эффекта.
Каждое местоположение функционально имитирует реакцию соответствующего класса активаторов (рис. 4B и 4D).Важно отметить, что стабильность состояния молчания снижается для третьего положения tetO по сравнению со вторым местом tetO. Это происходило даже в отсутствие этанола, что позволяет предположить, что разница в стабильности в молчаливом состоянии между активаторами класса II и Flo8p не является следствием альтернативного специфического для этанола эффекта.
Два режима репрессии Sfl1p соответствуют ступенчатому или неоднородному ответу
При выполнении титрования Sfl1p, описанного выше (рис. 4C и рис. 5A), мы сделали это на фоне sfl1Δ в этаноле. FLO11 был высоко экспрессирован (верхний левый квадрант) на фоне sfl1Δ , как было показано ранее [26]. Неожиданно оказалось, что при титровании Sfl1p в SD ura-media без этанола оба промотора выключались по шкале , класс (фиг. 5B). Это контрастировало с гетерогенной реакцией популяции, наблюдаемой при титровании активаторов класса II (фиг. 4), включая титрование Sfl1p в этаноле (фиг. 5A), при котором Flo8p предположительно более активен.Чтобы объяснить этот результат, мы предположили, что критический уровень Sfl1p необходим для замалчивания промотора, а ниже этого уровня Sfl1p все еще репрессирует транскрипцию, но обычным способом, связанным с более быстрыми колебаниями промотора. Модель требует, чтобы Sfl1p был способен репрессировать промотор FLO11 как глушитель или как обычный репрессор; доказательства существуют для обоих режимов [38], [40].
4), включая титрование Sfl1p в этаноле (фиг. 5A), при котором Flo8p предположительно более активен.Чтобы объяснить этот результат, мы предположили, что критический уровень Sfl1p необходим для замалчивания промотора, а ниже этого уровня Sfl1p все еще репрессирует транскрипцию, но обычным способом, связанным с более быстрыми колебаниями промотора. Модель требует, чтобы Sfl1p был способен репрессировать промотор FLO11 как глушитель или как обычный репрессор; доказательства существуют для обоих режимов [38], [40].
Рисунок 5. Hda1p необходим для подавления и гетерогенного ответа.
(A) Sfl1p, титрованный на фоне sfl1Δ в SD ура- + 1% этаноле, приводит к гетерогенному отклику.(B) Sfl1p, титрованный на фоне sfl1Δ в SD ura-, приводит к градиентному отклику. (C) Sfl1p, титрованный на фоне hda1Δ в SD ura- + 0,5% этаноле, возвращается к градиентному отклику. Экспрессия ниже без доксициклина из-за эндогенной экспрессии Sfl1p. (D) Анализ ChIP для выявления ацетилированных гистонов h4 и h5 на промоторе FLO11 (штаммы, выращенные в SD Complete или SD leu-). Зонды амплифицировали область промотора от -1,7 до -1,5 т.п.н. Сигнал (ось Y) представляет соотношение антиацетилированный гистон / антигистон или эффективное среднее ацетилирование на гистон в области.Удаление как sfl1 , так и hda1 приводит к гиперацетилированию промотора FLO11 , что связано с прекращением молчания. Следовательно, SFL1 -зависимое подавление звука на FLO11 требует HDA1 . Планки погрешностей представляют собой стандартную ошибку трехкратного количественного анализа образцов ПЦР. (E) Когда активатор Tec1p титруется на фоне hda1Δ , отклик также градуируется (SD ura-). (F) Средние уровни экспрессии во время титрования Sfl1p и Tec1p в фонах дикого типа (синяя кривая) и hda1Δ (розовая кривая).Устранение сайленсинга из-за недостатка Hda1p снижает пороговый уровень, при котором функционируют активаторы. Кроме того, оценивается реакция популяции (C и E, см. Рисунок S8 для титрования других активаторов).
Зонды амплифицировали область промотора от -1,7 до -1,5 т.п.н. Сигнал (ось Y) представляет соотношение антиацетилированный гистон / антигистон или эффективное среднее ацетилирование на гистон в области.Удаление как sfl1 , так и hda1 приводит к гиперацетилированию промотора FLO11 , что связано с прекращением молчания. Следовательно, SFL1 -зависимое подавление звука на FLO11 требует HDA1 . Планки погрешностей представляют собой стандартную ошибку трехкратного количественного анализа образцов ПЦР. (E) Когда активатор Tec1p титруется на фоне hda1Δ , отклик также градуируется (SD ura-). (F) Средние уровни экспрессии во время титрования Sfl1p и Tec1p в фонах дикого типа (синяя кривая) и hda1Δ (розовая кривая).Устранение сайленсинга из-за недостатка Hda1p снижает пороговый уровень, при котором функционируют активаторы. Кроме того, оценивается реакция популяции (C и E, см. Рисунок S8 для титрования других активаторов). Планки погрешностей представляют 3 s.d. около среднего, рассчитанного на основе анализа начальной загрузки. Вставка: Квадрат собственного шума Sfl1p (слева) и Tec1p (справа), титрованных в hda1Δ , пропорционален обратной величине содержания белка (здесь показан как средний уровень флуоресценции). Это указывает на то, что без молчания регуляторы модулируют экспрессию, контролируя частоту всплесков ( λ ‘).
Планки погрешностей представляют 3 s.d. около среднего, рассчитанного на основе анализа начальной загрузки. Вставка: Квадрат собственного шума Sfl1p (слева) и Tec1p (справа), титрованных в hda1Δ , пропорционален обратной величине содержания белка (здесь показан как средний уровень флуоресценции). Это указывает на то, что без молчания регуляторы модулируют экспрессию, контролируя частоту всплесков ( λ ‘).
https://doi.org/10.1371/journal.pgen.1000673.g005
Для проверки этой модели мы создали штамм с двумя репортерами на фоне hda1Δ и гетерологично добавили обратно Sfl1p. Когда мы титровали Sfl1p на этом фоне в SD ura-media, мы наблюдали постепенный ответ (рис. 5C). Это устанавливает, что Sfl1p способен репрессировать экспрессию Hda1p-независимым способом. Градиентный ответ предполагает, что Sfl1p работает как обычный репрессор, а не влияет на более медленные флуктуации между эпигенетически замалчиваемым и компетентным состояниями промотора. Чтобы дополнительно продемонстрировать, что Hda1p необходим, чтобы заглушить FLO11 , мы измерили среднее состояние ацетилирования гистонов h4 и h5 на промоторе FLO11 по хроматину-IP в штамме с двумя репортерами, выращенном в SD ura- (рис. 5D). Только промотор FLO11 дикого типа обнаруживает гипоацетилированное состояние, что указывает на замалчивание хроматина [41] и сходно с заглушенным теломерным участком. И в фонах hda1Δ и sfl1Δ промотор был гиперацетилирован, состояние хроматина ассоциировалось с отсутствием сайленсинга.Вместе это демонстрирует, что Sfl1p подавляет промотор Hda1p-зависимым образом, а состояние молчания на уровне FLO11 коррелирует с гипоацетилированием по крайней мере в одной области (~ -1600 п.н.) промотора.
Чтобы дополнительно продемонстрировать, что Hda1p необходим, чтобы заглушить FLO11 , мы измерили среднее состояние ацетилирования гистонов h4 и h5 на промоторе FLO11 по хроматину-IP в штамме с двумя репортерами, выращенном в SD ura- (рис. 5D). Только промотор FLO11 дикого типа обнаруживает гипоацетилированное состояние, что указывает на замалчивание хроматина [41] и сходно с заглушенным теломерным участком. И в фонах hda1Δ и sfl1Δ промотор был гиперацетилирован, состояние хроматина ассоциировалось с отсутствием сайленсинга.Вместе это демонстрирует, что Sfl1p подавляет промотор Hda1p-зависимым образом, а состояние молчания на уровне FLO11 коррелирует с гипоацетилированием по крайней мере в одной области (~ -1600 п.н.) промотора.
Если молчание устранено на фоне hda1Δ , титрование активаторов в присутствии высоких уровней Sfl1p приведет к постепенному ответу, поскольку Sfl1p теперь функционирует как обычный репрессор. Кроме того, пороговый уровень активатора, необходимый для включения FLO11 , будет ниже.Мы выполнили эти титрования (показано для Tec1p на фиг. 5E и 5F и других активаторов на фиг. S8) и подтвердили это предсказание. Кроме того, собственный шум этих штаммов был пропорционален обратному квадратному корню из среднего уровня экспрессии (вставка на рис. 5F и рис. S8), что указывает на то, что без медленных колебаний промотора активаторы регулируют частоту всплесков, λ ‘ [19 ].
Кроме того, пороговый уровень активатора, необходимый для включения FLO11 , будет ниже.Мы выполнили эти титрования (показано для Tec1p на фиг. 5E и 5F и других активаторов на фиг. S8) и подтвердили это предсказание. Кроме того, собственный шум этих штаммов был пропорционален обратному квадратному корню из среднего уровня экспрессии (вставка на рис. 5F и рис. S8), что указывает на то, что без медленных колебаний промотора активаторы регулируют частоту всплесков, λ ‘ [19 ].
Этанол контролирует важность подавления звука Flo8p-зависимым образом
Чтобы лучше понять роль этанола в передаче сигналов FLO11 , мы вырастили штамм с двумя репортерами в SD uramedia в диапазоне концентраций этанола.Экспрессия FLO11 проявляла дифференцированный ответ на повышение уровней этанола, но средний уровень экспрессии оставался низким даже при самых высоких (3%) концентрациях этанола (фиг. 6А). Градуированный ответ указывает на отсутствие сайленсинга, возможно, из-за повышенной активности Flo8p. Это привело к гипотезе, что при низких уровнях этанола (<1%) активность Flo8p устраняет опосредованное Sfl1p молчание, но мало влияет на экспрессию. Следовательно, хотя экспрессия FLO11 выключена (нижний правый квадрант), промотор фактически находится в компетентном состоянии.Чтобы проверить эту идею, мы титровали активаторы классов I и II в 1% этаноле. Оба класса были способны повышать экспрессию FLO11 и до высоких уровней способом с градацией (фиг. 6B), что означает, что в этих условиях молчание больше не происходило. Аналогичный эффект имели соответствующие синтетические активаторы. Напротив, титрование синтетического активатора, имитирующего Flo8p, привело к ответу, аналогичному этанолу (рис. 6C). Наконец, титрование активатора класса I и II в flo8Δ в SD ura- с 1% этанолом (рис. 6D) вернулось к поведению, наблюдаемому в SD ura- условиях.Следовательно, этанол контролирует степень сайленсинга на FLO11 Flo8p-зависимым образом. И Flo8p, и его синтетический аналог влияют на медленные переходы промотора, но ни один из них не является сильным традиционным активатором.
Это привело к гипотезе, что при низких уровнях этанола (<1%) активность Flo8p устраняет опосредованное Sfl1p молчание, но мало влияет на экспрессию. Следовательно, хотя экспрессия FLO11 выключена (нижний правый квадрант), промотор фактически находится в компетентном состоянии.Чтобы проверить эту идею, мы титровали активаторы классов I и II в 1% этаноле. Оба класса были способны повышать экспрессию FLO11 и до высоких уровней способом с градацией (фиг. 6B), что означает, что в этих условиях молчание больше не происходило. Аналогичный эффект имели соответствующие синтетические активаторы. Напротив, титрование синтетического активатора, имитирующего Flo8p, привело к ответу, аналогичному этанолу (рис. 6C). Наконец, титрование активатора класса I и II в flo8Δ в SD ura- с 1% этанолом (рис. 6D) вернулось к поведению, наблюдаемому в SD ura- условиях.Следовательно, этанол контролирует степень сайленсинга на FLO11 Flo8p-зависимым образом. И Flo8p, и его синтетический аналог влияют на медленные переходы промотора, но ни один из них не является сильным традиционным активатором.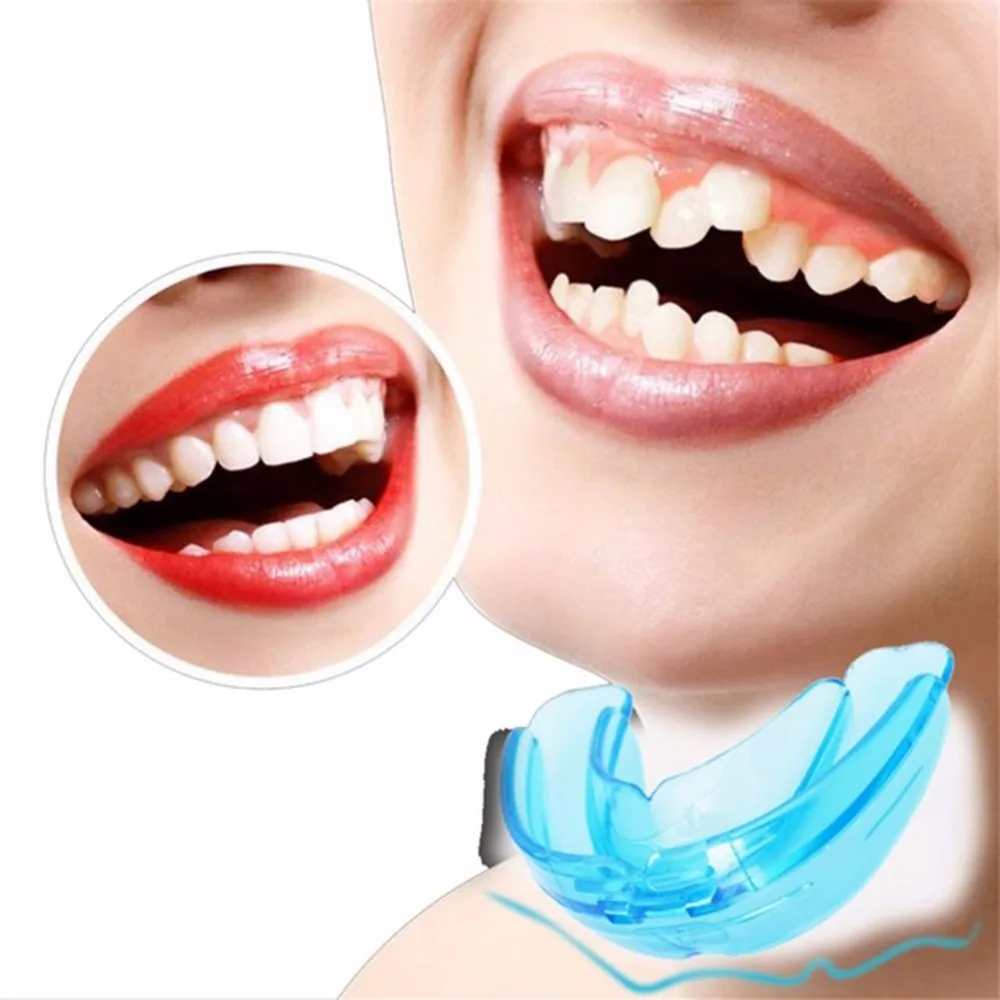
Рисунок 6. Этанол модулирует сайленсинг на промоторе через Flo8p.
(A) Дикий тип, выращенный в SD с добавлением этанола до конечной концентрации в диапазоне от 0 до 3%. (B) Активаторы, титрованные на фоне дикого типа в SD ura- + 1% этаноле. Все ответы оцениваются, что свидетельствует о потере молчания промотора.(C) Титрование синтетического активатора (rtTA) в SD ura- + 1% этаноле. Как и в (B), ответы также оцениваются. (D) Активаторы, титрованные в flo8Δ в SD ura- + 1% этаноле. Ответ ближе к тому, что представлен на рисунке 4A, а не на рисунке 6B, что указывает на то, что этанол устраняет молчание на промоторе через Flo8p. (E) Типичные гистограммы флуоресценции титров, показанные на (A, B, C, D). (Вверху) Wildtype, титрованный как на панели A, Tec1p, титрованный как на (B), и как на (D). (Внизу) rtTA, титрованная в штамме с сайтом tetO при -1160 (закрывающий сайт связывания Sfl1p) и при -350 (закрытый сайт нуклеосомы), как на панели C; Msn1p титруют как в (D).
https://doi.org/10.1371/journal.pgen.1000673.g006
Обсуждение
Основные результаты нашей работы, возможно, лучше всего можно понять с помощью модели с тремя состояниями на рисунке 7, простой, но полезной парадигмы для описания кинетики эпигенетической регуляции генов. Регуляторы могут влиять либо на медленные скорости перехода, связанные с эпигенетическим молчанием, либо на быстрые скорости переходов, связанные с обычной активацией генов. Хотя о том, как регуляторы влияют на быстрые скорости перехода, известно много, меньше известно о том, как регуляторы влияют на медленные скорости перехода и связаны ли эффекты на медленные и быстрые скорости перехода.Наша работа демонстрирует, что медленные переходы у двух копий промотора FLO11 в диплоидных дрожжах происходят случайно и независимо. Кроме того, мы определяем роль различных регуляторов trans из FLO11 в управлении как медленной, так и быстрой скоростью перехода; похоже, что этот контроль распределен между различными «классами» регуляторов. Важно отметить, что распределенный контроль позволяет клетке формировать разнообразие экспрессии FLO11 в изогенной популяции за счет использования различных комбинаций регуляторов.
Важно отметить, что распределенный контроль позволяет клетке формировать разнообразие экспрессии FLO11 в изогенной популяции за счет использования различных комбинаций регуляторов.
Рисунок 7. Функциональные роли регуляторов активации промотора FLO11 формируют ответ популяции.
Молчание на промоторе устанавливается связыванием Sfl1p и привлечением Hda1p. Относительные активности Flo8p и Sfl1p определяют состояние хроматина промотора. Основное состояние промотора OFF-популяции может быть выявлено путем добавления активаторов класса I и II (внизу), поскольку активаторы класса I не могут эффективно активировать транскрипцию на молчащем промоторе, тогда как активаторы класса II могут активировать экспрессию, достаточно стабилизируя компетентное состояние.На очень высоких уровнях активаторы класса II нарушают молчание; в этот момент все клетки также имеют высокую экспрессию. Напротив, промежуточный уровень активности Flo8p может «открыть» промотор, переводя молчание в стабильное компетентное состояние, в то время как экспрессия остается низкой.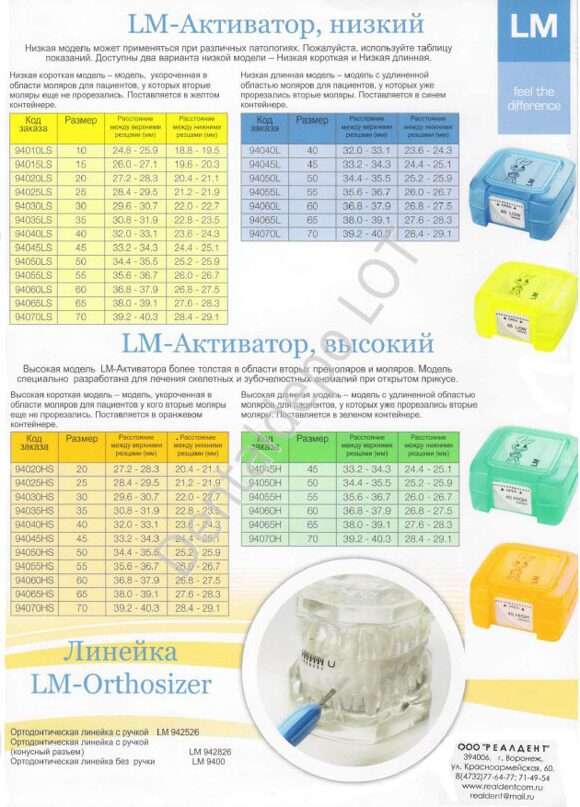 Это открытие может быть связано с модификациями хроматина. Комбинация активации Flo8p и активаторов класса I позволяет разделить состояние хроматина и уровень экспрессии, тогда как активация только активаторами класса II этого не позволяет.
Это открытие может быть связано с модификациями хроматина. Комбинация активации Flo8p и активаторов класса I позволяет разделить состояние хроматина и уровень экспрессии, тогда как активация только активаторами класса II этого не позволяет.
https://doi.org/10.1371/journal.pgen.1000673.g007
Наша демонстрация того, что две копии промотора FLO11 переключаются медленно и независимо, основывается на предыдущей работе, демонстрирующей (1), что промотор FLO11 регулируется эпигенетически. [30] и (2) две копии частично заглушенного промотора URA3 , вставленного в локусы типа спаривания в S. cerevisiae , переключаются независимо на фоне sir1 [24]. Теперь мы предоставляем доказательства того, что независимое переключение происходит в естественном гене, эпигенетическая регуляция которого не зависит от белка SIR.Учитывая богатое разнообразие генофонда FLO11 и [29], независимое переключение может быть дополнительным механизмом для генерации вариации адгезивного фенотипа.
Чтобы понять, как контролируется гетерогенность экспрессии на FLO11 , мы взяли функциональный подход и определили кинетическую роль различных регуляторов как в медленных, так и в быстрых колебаниях промотора. Класс I регулирует исключительно быстрые колебания промотора. Измерения внутреннего шума подтвердили, что эти регуляторы дестабилизируют компетентное состояние и увеличивают частоту всплесков, что является общей темой для регуляторов дрожжей [19], [20].Интересно, что мы обнаружили, что Class II и Flo8p регулируют медленные колебания промотора в первую очередь за счет стабилизации компетентного состояния. Предыдущее исследование локусов типа спаривания обнаружило, что активатор, Ppr1p, может вызывать молчание и влиять на обе скорости медленных переходов [24], но это было измерено только на одном уровне Ppr1p. Только путем титрования регуляторов мы смогли четко определить их функциональные роли. Влияют ли активаторы в целом на одну или обе скорости перехода молчащих генов, остается открытым вопросом, но, вероятно, зависит от механизма молчания.
В то время как титрование регулятора показывает, что этанол обессиливает промотор через Flo8p, путь (пути), с помощью которого этанол активирует Flo8p, неизвестен. Самый простой механизм заключается в том, что длительный рост в этаноле активирует PKA (в частности, Tpk2p), который затем активирует Flo8p и деактивирует Sfl1p. Фактически, глюкоза не требуется как промоторный ответ, когда Flo8p титровали в синтетической среде этанолом или глицерином, поскольку единственный источник углерода был аналогичен таковому в SD 1% этаноле (данные не показаны).Однако об активации пути PKA этанолом не сообщалось. Также возможно, что Flo8p активируется этанолом через другой путь, независимый от PKA, хотя, насколько нам известно, он не является мишенью для какой-либо другой киназы. Третья возможность заключается в том, что этанол может обеспечивать синергетические взаимодействия между Flo8p и другими регуляторами, что приводит к десиленсированию, хотя нет никаких доказательств взаимодействия Flo8p с какими-либо активаторами класса I. Эти возможности можно выявить путем мониторинга уровней циклического АМФ и статуса фосфорилирования Flo8p и Sfl1p в этаноле.
Эти возможности можно выявить путем мониторинга уровней циклического АМФ и статуса фосфорилирования Flo8p и Sfl1p в этаноле.
Наши результаты подтверждают идею, что положение сайта связывания в промоторе FLO11 может во многом определять кинетическую роль регулятора транскрипции. Однако механистическое описание того, как связывание с конкретными сайтами влияет на медленную эпигенетическую регуляцию и быструю традиционную регуляцию, а также молекулярная природа молчаливых и компетентных состояний промотора все еще остается неясным. Механистическое объяснение двойной роли Sfl1p, вероятно, является наиболее ясным. Связывание в области, свободной от -1200 нуклеосом, регулирует эпигенетическое молчание, возможно, за счет рекрутирования Hda1p через корепрессор Tup1p / Ssn6p [38].Обычная регуляция, скорее всего, происходит посредством связывания Sfl1p с областью -200, которая содержит предполагаемый сайт связывания Sfl1p и, как было показано, связывает Sfl1p in vitro . В самом деле, предварительные эксперименты с ChIP предполагают, что Sfl1p связан с этой областью in vivo (Octavio и Maheshri, неопубликованные результаты).
В самом деле, предварительные эксперименты с ChIP предполагают, что Sfl1p связан с этой областью in vivo (Octavio и Maheshri, неопубликованные результаты).
Среди активаторов роль Flo8p и его синтетического аналога, пожалуй, наиболее очевидна. Связывание Flo8p и связывание Sfl1p в области -1200, вероятно, являются взаимоисключающими из-за перекрывающихся сайтов связывания, и поэтому Flo8p может предотвращать Sfl1p-опосредованное установление сайленсинга, но, вероятно, не влияет напрямую на обычную репрессию Sfl1p.Это могло бы объяснить способность Flo8p и его соответствующего синтетического аналога влиять на медленный эпигенетический переход независимо от обычной активации. Фактически, связывание tetR, в котором отсутствует домен активации VP16 rtTA, с сайтом -1160 tetO достаточно для нарушения сайленсинга (данные не показаны), подразумевая, что стерические препятствия являются основным способом действия. Более того, любая слабая активация через Flo8p может быть связана с его известной ролью в связывании с промоторами других активаторов класса I (включая те, что были протестированы здесь) и, предположительно, в повышении их экспрессии. Таким образом, активация Flo8p может привести промотор FLO11 и в компетентное, «уравновешенное» состояние, экспрессия которого может контролироваться регуляторами класса I. Однако есть свидетельства того, что Flo8p может связываться с другими участками промотора [31].
Таким образом, активация Flo8p может привести промотор FLO11 и в компетентное, «уравновешенное» состояние, экспрессия которого может контролироваться регуляторами класса I. Однако есть свидетельства того, что Flo8p может связываться с другими участками промотора [31].
Существует несколько возможностей неспособности активаторов класса I вызывать молчание, но все же регулировать частоту пакетов компетентного промотора FLO11 . Например, близость сайта связывания к сайту начала транскрипции может играть доминирующую роль.Канонические дрожжевые промоторы обычно имеют размер 150–400 п.н., а сайты связывания транскрипционных факторов сгруппированы в 100–200 п.н. от сайта начала транскрипции [42]; эта близость позволяет напрямую взаимодействовать с общим транскрипционным аппаратом. Следовательно, возможно, что активаторы класса I связываются в центральной области промотора FLO11 , области, которая может быть недоступна для факторов транскрипции и / или транскрипционного аппарата в состоянии молчания. Однако, в то время как сайт связывания синтетического аналога класса I находится на -350 в области, закрытой нуклеосомой, не все активаторы класса I имеют сайты связывания в этой области [31].
Однако, в то время как сайт связывания синтетического аналога класса I находится на -350 в области, закрытой нуклеосомой, не все активаторы класса I имеют сайты связывания в этой области [31].
Вторая возможность, не исключающая друг друга с первой, состоит в том, что активаторам класса I не нужно связываться в нижележащей области, но они могут влиять на скорость транскрипции с помощью механизмов дальнего действия (но быстрых), включая образование петель ДНК, криптическую транскрипцию или ремоделирование хроматина дальнего действия. Это могло бы объяснить присутствие сайтов связывания активатора класса I в этих областях, которые, как известно, связаны in vivo с в активирующих условиях [31]. Кроме того, это может объяснить, почему даже при высоких уровнях экспрессии в отсутствие сайленсинга внутренний шум на промоторе FLO11 в 10 раз выше, чем у аналогичного высоко экспрессируемого промотора PHO84 (данные не показаны).Однако, при любом объяснении, неспособность активаторов класса I бросить вызов состоянию молчания неясна. Измененная структура хроматина или сниженная доступность сайтов связывания могут быть вызваны тем, что сайты активаторов класса I, как правило, находятся под нуклеосомами. Однако, за исключением некоторого истощения нуклеосом в коровом промоторе и области -1300, грубые нуклеосомные перестройки, по-видимому, не происходят при молчании (Рисунок S9), хотя картирование с более высоким разрешением может выявить более тонкие различия.
Измененная структура хроматина или сниженная доступность сайтов связывания могут быть вызваны тем, что сайты активаторов класса I, как правило, находятся под нуклеосомами. Однако, за исключением некоторого истощения нуклеосом в коровом промоторе и области -1300, грубые нуклеосомные перестройки, по-видимому, не происходят при молчании (Рисунок S9), хотя картирование с более высоким разрешением может выявить более тонкие различия.
В то время как мы не знаем биохимических промежуточных продуктов во время медленных переходов промоторов, скорости псевдопервого порядка предполагают одиночный медленный шаг, а не распределенный механизм контроля.Это похоже как на локусы с частично подавленным типом спаривания [24], так и на эпигенетически регулируемый ген agn43 в E. coli [43]. Возможности для медленного эпигенетического шага, управляющего от ON к OFF, могут включать, среди прочего, связывание Sfl1p или опосредованное Sfl1p рекрутирование факторов сайленсинга. Оба активатора класса II Msn1p и Mss11p способны стабилизировать активное состояние, но их локализация и активность по отношению к промотору FLO11 остается неясной [32], [34].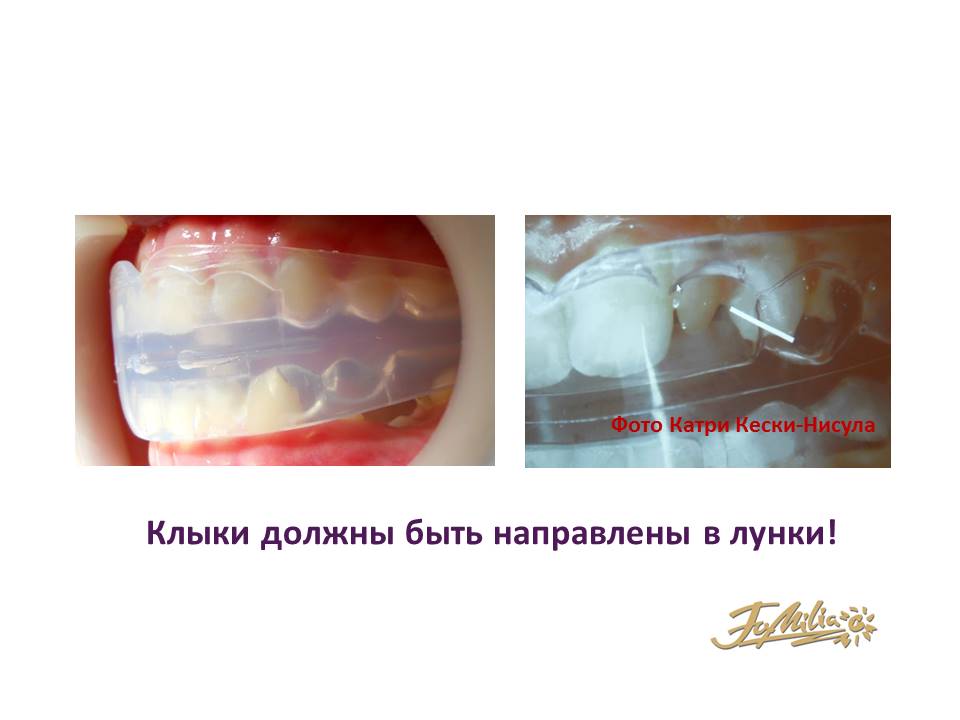 Способность синтетического аналога класса II оказывать аналогичное стабилизирующее действие на состояние ON за счет связывания с областью -1470, а также состояние дифференциального ацетилирования этой области настоятельно предполагает ремоделирование хроматина в области выше по течению, что влияет на доступность активаторов класса I и переход в дееспособное состояние. Картирование с высоким разрешением состояния хроматина всего промотора FLO11 в различных условиях должно указывать на биохимический механизм медленного перехода промотора и будет в центре внимания будущей работы.
Способность синтетического аналога класса II оказывать аналогичное стабилизирующее действие на состояние ON за счет связывания с областью -1470, а также состояние дифференциального ацетилирования этой области настоятельно предполагает ремоделирование хроматина в области выше по течению, что влияет на доступность активаторов класса I и переход в дееспособное состояние. Картирование с высоким разрешением состояния хроматина всего промотора FLO11 в различных условиях должно указывать на биохимический механизм медленного перехода промотора и будет в центре внимания будущей работы.
Наши результаты имеют значение для регуляции различных субтеломерно кодируемых семейств генов, которые, как известно, регулируются эпигенетически. Это включает семейство генов FLO [30] и другие близкородственные адгезины дрожжей [25], такие как семейство генов EPA в патогенных дрожжах C. glabrata [44]. Фенотипическая изменчивость в экспрессии гена EPA может позволить C. glabrata быстро колонизировать новые ткани хозяина и избежать иммунного надзора.Включаются и выключаются такие гены независимо, зависит ли это от механизмов их сайленсинга (SIR-зависимых и т. Д.), Их относительного положения в хромосомах или наличия граничных элементов? Какие промоторные переходы регулируют факторы транс и ? Это понимание позволит конструировать штаммы с четко определенными уровнями фенотипической гетерогенности. Такие штаммы являются предпосылкой для количественной оценки того, какую роль, если какая-либо фенотипическая гетерогенность играет на приспособленности организма в естественной среде.
glabrata быстро колонизировать новые ткани хозяина и избежать иммунного надзора.Включаются и выключаются такие гены независимо, зависит ли это от механизмов их сайленсинга (SIR-зависимых и т. Д.), Их относительного положения в хромосомах или наличия граничных элементов? Какие промоторные переходы регулируют факторы транс и ? Это понимание позволит конструировать штаммы с четко определенными уровнями фенотипической гетерогенности. Такие штаммы являются предпосылкой для количественной оценки того, какую роль, если какая-либо фенотипическая гетерогенность играет на приспособленности организма в естественной среде.
Материалы и методы
Штаммы дрожжей и среды
Чтобы использовать анализ с двумя репортерами для изучения переключения промотора FLO11 , мы заменили ORF FLO11 в гаплоиде Σ1278b из лаборатории Хейтмана [45] на Y FP- K и MX6 или CFP-K. Кассеты и MX6 интеграцией ПЦР, а затем спаривание для создания диплоидов. Все использованные штаммы и плазмиды представлены в Таблице S1 и Таблице S2.
Все использованные штаммы и плазмиды представлены в Таблице S1 и Таблице S2.
SD — синтетическая среда с 2% глюкозы.В SD ura-media отсутствует урацил. SD ura-media с этанолом содержит как 2% глюкозы, так и определенное количество этанола.
Измерения и анализ отдельных ячеек
Клетки из ночных культур, выращенных в этаноле SD ura- или SD ura- +, инокулировали при исходной OD 600 от 0,005 до 0,01 и выращивали в течение 15-20 часов в той же среде. Для экспериментов по титрованию эти культуры обрабатывали серийными разведениями доксицилина (от 0 до 10000 нг / мл) при 30 ° C в хорошо встряхиваемых 96-луночных планшетах с глубокими лунками.Клетки собирали в средне-поздней логарифмической фазе (OD 600 от 0,5 до 1,5) и помещали на лед, пока обрабатывались другие образцы. Экспрессию измеряли с помощью микроскопа Zeiss AxioObserver с фильтрами, оптимизированными для yECitrine, mCherry и Cerulean (Chroma). Программное обеспечение Metamorph (Molecular Devices) использовалось для анализа изображений и количественной оценки флуоресценции YFP и CFP отдельных клеток. Для каждого образца было отображено от 500 до 1500 клеток. Уровни флуоресценции в канале RFP использовали для удаления мертвых клеток (обычно <5% популяции).Подробности предварительной обработки данных и оценки λ и γ приведены в Тексте S1.
Для каждого образца было отображено от 500 до 1500 клеток. Уровни флуоресценции в канале RFP использовали для удаления мертвых клеток (обычно <5% популяции).Подробности предварительной обработки данных и оценки λ и γ приведены в Тексте S1.
В экспериментах по таймлапс-микроскопии клетки загружали на микрожидкостную платформу ONIX (CellASIC) с первоначально ~ 10 клетками, захваченными в отдельные камеры. Среда в камерах ∼10 -5 мкл постоянно пополнялась со скоростью 10 мкл / час. YFP, CFP, RFP флуоресценция и изображения в светлом поле получали каждые 15 минут в течение 20 часов. Пример представлен в Video S1.Стеки изображений были сегментированы с использованием настраиваемых журналов Metamorph. Отслеживание отдельных клеток и флуоресценцию определяли с использованием пользовательских процедур MATLAB.
Анализ иммунопреципитации хроматина
ИП хроматина были выполнены по методике [46]. Вкратце, ~ 40 мл клеток выращивали при 30 ° C либо в SD полной, либо в SD лей- до OD 600 = 0,8. Лизаты фиксированных клеток обрабатывали ультразвуком для срезания хроматина до средней длины 500 п.н., а выделенный хроматин инкубировали с 2 мкл антител (Upstate / Millipore) против любого гистона h4 (кат.No. 05-928), гистон h5 (Cat. No. 05-858), ацетилированный гистон h4 (Cat. No. 07-593) и ацетилированный гистон h5 (Cat. No. 06-866). Образец без антител также был приготовлен в качестве контроля. После обращения поперечных сшивок ДНК из хроматина, подвергнутого иммунопреципитации, очищали и анализировали с помощью количественной ПЦР (Applied Biosystems). Использовали праймеры, амплифицирующие область от -1,7 до -1,5 т.п.н. промотора FLO11 , и праймеры, амплифицирующие область теломера в правом плече хромосомы VI [47] в качестве контроля сигналов гипоацетилированных гистонов.Программное обеспечение Applied Biosystems 7300 использовалось для получения пороговых значений цикла. Все сигналы от экспериментальных образцов были количественно определены относительно сигнала от известного количества геномной ДНК от немодифицированного когенного штамма Σ1287b (MLY43, Таблица S1), который служил в качестве положительного контроля.
Лизаты фиксированных клеток обрабатывали ультразвуком для срезания хроматина до средней длины 500 п.н., а выделенный хроматин инкубировали с 2 мкл антител (Upstate / Millipore) против любого гистона h4 (кат.No. 05-928), гистон h5 (Cat. No. 05-858), ацетилированный гистон h4 (Cat. No. 07-593) и ацетилированный гистон h5 (Cat. No. 06-866). Образец без антител также был приготовлен в качестве контроля. После обращения поперечных сшивок ДНК из хроматина, подвергнутого иммунопреципитации, очищали и анализировали с помощью количественной ПЦР (Applied Biosystems). Использовали праймеры, амплифицирующие область от -1,7 до -1,5 т.п.н. промотора FLO11 , и праймеры, амплифицирующие область теломера в правом плече хромосомы VI [47] в качестве контроля сигналов гипоацетилированных гистонов.Программное обеспечение Applied Biosystems 7300 использовалось для получения пороговых значений цикла. Все сигналы от экспериментальных образцов были количественно определены относительно сигнала от известного количества геномной ДНК от немодифицированного когенного штамма Σ1287b (MLY43, Таблица S1), который служил в качестве положительного контроля. Отношение сигналов антигистоновых / антиацетилированных гистонов использовали в качестве меры среднего ацетилирования h4 и h5 в области.
Отношение сигналов антигистоновых / антиацетилированных гистонов использовали в качестве меры среднего ацетилирования h4 и h5 в области.
Анализ на микрококковую нуклеазу
Тесты на микрококковую нуклеазу выполняли, как в [48].Все клетки выращивали при 30 ° C либо в SD полной, либо в SD лей- до OD 600 = 0,5.
Дополнительная информация
Рисунок S1.
Данные о занятости нуклеосом из рисунка S9 нанесены на график вместе с предсказанными положениями нуклеосом из исследований, предсказывающих положение нуклеосом для всего генома. Исследование Kaplan et al [17] обеспечивает расчетное предсказание положения нуклеосом в любой последовательности. Прогнозируется только несколько бедных нуклеосомами областей, граничащих с четко определенными нуклеосомами, включая область -1200.В исследовании Mavrich et al [18] использовалась статистическая модель для анализа экспериментальных данных о нуклеосомной занятости всего генома. Столбцы x-ошибок обозначают, насколько нечетким является положение. Относительная шкала занятости является произвольной, и абсолютное число нельзя сравнивать между наборами данных. Однако некоторые общие тенденции в позиционированных и нечетких нуклеосомах очевидны, и это было основой рисунка 1 в основном тексте.
Относительная шкала занятости является произвольной, и абсолютное число нельзя сравнивать между наборами данных. Однако некоторые общие тенденции в позиционированных и нечетких нуклеосомах очевидны, и это было основой рисунка 1 в основном тексте.
https://doi.org/10.1371/journal.pgen.1000673.s001
(0,46 МБ TIF)
Рисунок S2.
Экспрессия FLO11 на твердой среде — одиночные репортерные штаммы. 10 мкл средне-логарифмической культуры различных диплоидных штаммов с отдельными репортерами наносили на свежий планшет YPD. Метки слева показывают, была ли и в каком локусе ORF FLO11 заменена конкретным вариантом флуоресцентного белка. Планшеты оставляли при комнатной температуре. Были взяты образцы клеток из всех областей пятна (см. Текст S1), и экспрессия CFP и YFP в этих образцах контролировалась каждые 2 дня с помощью флуоресцентной микроскопии.Приведены графики плотности для каждого образца, где по оси абсцисс отложены уровни флуоресценции log CFP, а по оси y — уровни флуоресценции log YFP. Аутофлуоресценцию клеток можно оценить на основе (первого) контрольного штамма. Оба репортера флуоресценции реагируют одинаково, независимо от того, интегрированы ли они в локус A или α.
Аутофлуоресценцию клеток можно оценить на основе (первого) контрольного штамма. Оба репортера флуоресценции реагируют одинаково, независимо от того, интегрированы ли они в локус A или α.
https://doi.org/10.1371/journal.pgen.1000673.s002
(1,26 МБ TIF)
Рисунок S3.
Экспрессия FLO11 на твердой среде — двойные репортерные штаммы.Как на панели Рисунок S2. Были сконструированы четыре различных двойных репортерных штамма, два с CFP в α-локусе и YFP в A-локусе и два в противоположной конфигурации. Их ответ похож, подтверждая, что оба репортера равнозначны. Кроме того, распределение экспрессии каждого отдельного флуоресцентного репортера эквивалентно соответствующему отдельному репортерному штамму на рисунке S2, что подтверждает независимость и тот факт, что экспрессия FLO11 не имеет обратной связи и не влияет на свою собственную экспрессию.
https://doi.org/10.1371/journal.pgen.1000673.s003
(1,28 МБ TIF)
Рисунок S4.

Статические снимки клеток Y45 в YP 1% этанола, 2% глицерина, поддерживаемых в экспоненциальной фазе разбавлением. Нулевая гипотеза о том, что распределения флуоресценции YFP в каждый момент времени эквивалентны, не может быть отвергнута (двусторонний тест Колмогорова-Смирнова, p = 0,60, 0,25, 0,85 для дня 1 по сравнению с днем 2, день 2 по сравнению с днем 3 и день 1 по сравнению с 3-м днем соответственно). Точно так же нельзя отвергнуть нулевую гипотезу об эквивалентности распределений флуоресценции CFP в каждый момент времени (двусторонний тест Колмогорова-Смирнова, p = 0.66, 0,36, 0,88 для дня 1 против дня 2, дня 2 против дня 3 и дня 1 против дня 3 соответственно).
https://doi.org/10.1371/journal.pgen.1000673.s004
(0,45 МБ TIF)
Рисунок S6.
Скорость переключения репортера CFP. Как на рисунке 3A в основном тексте, но для CFP, а не для YFP. Подгонка дает скорости переключения для CFP: λ / δ (ВЫКЛ-ВКЛ) = 0,25 + 0,03 поколение -1 (розовый), γ / δ (ВКЛ-ВЫКЛ) = 0,90 + 0,17 поколение -1 (синий). Планки погрешностей соответствуют 3 s.d. от среднего, рассчитанного с помощью бутстрап-анализа.
Планки погрешностей соответствуют 3 s.d. от среднего, рассчитанного с помощью бутстрап-анализа.
https://doi.org/10.1371/journal.pgen.1000673.s006
(0,64 МБ TIF)
Рисунок S7.
Переключатель ВЫКЛ (ВКЛ) -ВКЛ (ВЫКЛ) на FLO11 не коррелирует со стадией клеточного цикла. Время, когда рожденная клетка ВЫКЛЮЧАЕТСЯ, включается (A) или время, когда рожденная клетка выключается (В) после того, как произошло ее самое последнее событие деления, показано для каждой клетки, переключившейся во время эксперимента с интервальной съемкой, на рисунке 3А.Точки, в которых происходит переключение, по-видимому, не группируются в какой-либо конкретной позиции во время клеточного цикла.
https://doi.org/10.1371/journal.pgen.1000673.s007
(1,23 МБ TIF)
Рисунок S8.
Потеря Hda1p превращает гетерогенный промоторный ответ на активаторы Mss11p и Phd1p в дифференцированный ответ. Как показано на рисунке 4B и рисунке 5E в основном тексте, где Tec1p был титрован на фоне дикого типа (Y45) и hda1Δ соответственно, другие активаторы Phd1p и Mss11p также демонстрируют дифференцированный ответ на фоне hda1Δ (A и B). .На фоне дикого типа Phd1p (C), такой как Tec1p, неспособен стабилизировать состояние ON в достаточной степени, чтобы войти в бимодальный режим, тогда как Mss11p (D), как Msn1p (показанный на рисунке 4B), способен это сделать. Как показано на фиг. 5F, устранение молчания на фоне hda1Δ снижает пороговый уровень, при котором функционируют Mss11p и Phd1p (E и F). Планки погрешностей представляют собой 3 стандартных отклонения от среднего значения из начального анализа. Как и Tec1p в hda1Δ на рисунке 5F, и Mss11p, и Phd1p контролируют частоту всплеска ( λ ′) в отсутствие глушителя, как квадрат собственного шума Phd1p, титрованного в hda1Δ (G) и Mss11p, титрованного в hda1Δ (H) шкала с обратной величиной содержания белка.Все титрования проводили в SD ura-.
.На фоне дикого типа Phd1p (C), такой как Tec1p, неспособен стабилизировать состояние ON в достаточной степени, чтобы войти в бимодальный режим, тогда как Mss11p (D), как Msn1p (показанный на рисунке 4B), способен это сделать. Как показано на фиг. 5F, устранение молчания на фоне hda1Δ снижает пороговый уровень, при котором функционируют Mss11p и Phd1p (E и F). Планки погрешностей представляют собой 3 стандартных отклонения от среднего значения из начального анализа. Как и Tec1p в hda1Δ на рисунке 5F, и Mss11p, и Phd1p контролируют частоту всплеска ( λ ′) в отсутствие глушителя, как квадрат собственного шума Phd1p, титрованного в hda1Δ (G) и Mss11p, титрованного в hda1Δ (H) шкала с обратной величиной содержания белка.Все титрования проводили в SD ura-.
https://doi.org/10.1371/journal.pgen.1000673.s008
(3,07 МБ TIF)
Рисунок S9.
Микрококковое картирование нуклеазой FLO11 было выполнено на клетках, выращенных в условиях, когда промотор полностью подавлен (рост Y45 дикого типа в SD завершен) в (A) или полностью активен (рост штамма sfl1Δ в SD полная плюс 2% глюкозы) в (B). Общая структура хорошо согласуется с предсказаниями in silico на Рисунке 1.Хотя нуклеосомная занятость в области -1300 п.н. и области -150 п.н., по-видимому, еще больше истощается в активном состоянии, неожиданно нет грубой перестройки нуклеосомной структуры между молчащим и активным состояниями. Планки погрешностей представляют собой стандартную ошибку для трех количественных образцов ПЦР.
Общая структура хорошо согласуется с предсказаниями in silico на Рисунке 1.Хотя нуклеосомная занятость в области -1300 п.н. и области -150 п.н., по-видимому, еще больше истощается в активном состоянии, неожиданно нет грубой перестройки нуклеосомной структуры между молчащим и активным состояниями. Планки погрешностей представляют собой стандартную ошибку для трех количественных образцов ПЦР.
https://doi.org/10.1371/journal.pgen.1000673.s009
(0,55 МБ TIF)
Видео S1.
Фильм о росте клеток в микрофлюидном устройстве. Красный и зеленый представляют экспрессию CFP и YFP соответственно.В этом видео легко увидеть переходы из ВКЛ в ВЫКЛ и из ВЫКЛ в ВКЛ для каждой копии промоутера FLO11 . Каждый кадр представляет прошедшие 15 минут, а весь фильм представляет собой 20 часов роста.
https://doi.org/10.1371/journal.pgen.1000673.s013
(3,83 МБ AVI)
Благодарности
Мы благодарим Э. К. О’Ши и К. Верстрепена за ценные предложения и замечания; Б. Ниснер и Т. Л. К для штаммов и плазмид; C.J. Zopf за скрипты, используемые для таймлапс-анализа; К.Соломону и К. Л. Дж. Пратеру за техническую помощь; и С. Бумгарнеру, Г. Финку, А. Раджу, С. Рагхавану и М. Л. Онгу за критическое прочтение рукописи.
Ниснер и Т. Л. К для штаммов и плазмид; C.J. Zopf за скрипты, используемые для таймлапс-анализа; К.Соломону и К. Л. Дж. Пратеру за техническую помощь; и С. Бумгарнеру, Г. Финку, А. Раджу, С. Рагхавану и М. Л. Онгу за критическое прочтение рукописи.
Вклад авторов
Задумал и спроектировал эксперименты: LO NM. Проведены эксперименты: ЛО КГ НМ. Проанализированы данные: ЛО НМ. Предоставленные реагенты / материалы / инструменты анализа: LO NM. Написал статью: ЛО НМ.
Ссылки
- 1. Рандо О.Дж., Верстрепен К.Дж. (2007) Временные рамки генетического и эпигенетического наследования.Ячейка 128 (4): 655–668.
- 2. Эйвери С.В. (2006) Индивидуальность микробных клеток и основные источники неоднородности. Nat Rev Microbiol 4 (8): 577–587.
- 3. Barry JD, Ginger ML, Burton P, McCulloch R (2003) Почему гены контингентности паразитов часто связаны с теломерами? Int J Parasitol 33 (1): 29–45.
- 4.
Pays E, Vanhamme L, Perez-Morga D (2004) Антигенная изменчивость трипаносомы brucei: факты, проблемы и загадки. Curr Opin Microbiol 7 (4): 369–374.
- 5. Low DA, Weyand NJ, Mahan MJ (2001) Роль метилирования аденина ДНК в регуляции экспрессии и вирулентности бактериальных генов. Заражение иммунной 69 (12): 7197–7204.
- 6. Wu J, Grunstein M (2000) через 25 лет после нуклеосомной модели: модификации хроматина. Тенденции Biochem Sci 25 (12): 619–623.
- 7. Кауфманн BB, Ян К., Меттетал Дж. Т., ван Ауденаарден А. (2007) Наследственное стохастическое переключение, выявленное генеалогией одной клетки.PLoS Biol 5 (9): e239.
- 8. Gardner TS, Cantor CR, Collins JJ (2000) Конструирование генетического тумблера в Escherichia coli. Nature 403 (6767): 339–342.
- 9. Xiong W, Ferrell JE Jr (2003) Основанный на положительной обратной связи бистабильный «модуль памяти», который управляет решением судьбы клетки. Nature 426 (6965): 460–465.
- 10.
Hernday AD, Braaten BA, Broitman-Maduro G, Engelberts P, Low DA (2004) Регулирование эпигенетического переключения pap с помощью CpxAR: фосфорилированный CpxR ингибирует переход в состояние включения фазы за счет конкуренции с lrp. Молекулярная ячейка 16 (4): 537–547.
- 11. Golding I, Paulsson J, Zawilski SM, Cox EC (2005) Кинетика активности генов у отдельных бактерий в реальном времени. Ячейка 123 (6): 1025–1036.
- 12. Cai L, Friedman N, Xie XS (2006) Стохастическая экспрессия белка в отдельных клетках на уровне одной молекулы. Nature 440 (7082): 358–362.
- 13. Chubb JR, Trcek T, Shenoy SM, Singer RH (2006) Транскрипционная пульсация гена развития. Текущая биология 16 (10): 1018–1025.
- 14. Yu J, Xiao J, Ren X, Lao K, Xie XS (2006) Исследование экспрессии генов в живых клетках, по одной молекуле белка за раз. Science 311 (5767): 1600–1603.
- 15. Блейк В.Дж., Балажи Г., Кохански М.А., Айзекс Ф.Дж., Мерфи К.Ф. и др. (2006) Фенотипические последствия опосредованного промотором транскрипционного шума. Mol Cell 24 (6): 853–865.
- 16. Kaern M, Elston TC, Blake WJ, Collins JJ (2005) Стохастичность в экспрессии генов: от теорий до фенотипов.Nat Rev Genet 6 (6): 451–64.
- 17. Махешри Н., О’Ши Е.К. (2007) Жизнь с шумными генами: как надежно функционируют клетки с присущей им вариабельностью экспрессии генов. Анну Рев Биофиз Биомол Структура 36: 413–434.
- 18. Elowitz MB, Levine AJ, Siggia ED, Swain PS (2002) Стохастическая экспрессия гена в одной клетке. Наука 297 (5584): 1183
- 19. Бар-Эвен А., Полссон Дж., Махешри Н., Карми М., О’ши Е. и др. (2006) Уровень шума в экспрессии белка зависит от естественного содержания белка.Нат Генет 38 (6): 636–643.
- 20. Newman JR, Ghaemmaghami S, Ihmels J, Breslow DK, Noble M и др. (2006) Одноклеточный протеомный анализ S. cerevisiae раскрывает архитектуру биологического шума. Nature 441 (7095): 840–846.
- 21. Raser JM, O’Shea EK (2004) Контроль стохастичности в экспрессии эукариотических генов. Science 304 (5678): 1811–1814.
- 22. Mettetal JT, Muzzey D, Pedraza JM, Ozbudak EM, van Oudenaarden A (2006) Прогнозирование динамики экспрессии стохастических генов в отдельных клетках.PNAS 103 (19): 7304–7309.
- 23. Ingolia NT, Murray AW (2007) Петли положительной обратной связи как гибкий биологический модуль. Текущая биология 17 (8): 668–677.
- 24. Xu EY, Zawadzki KA, Broach JR (2006) Одноклеточные наблюдения выявляют промежуточные состояния транскрипционного молчания. Mol Cell 23 (2): 219–229.
- 25. Verstrepen KJ, Klis FM (2006) Флокуляция, адгезия и образование биопленок у дрожжей. Мол микробиол 60 (1): 5–15.
- 26.Rupp S, Summers E, Lo HJ, Madhani H, Fink G (1999) MAP-киназа и сигнальные пути филаментации цАМФ сходятся на необычно большом промоторе дрожжевого гена FLO11. EMBO J 18 (5): 1257–1269.
- 27. Pan X, Heitman J (2002) Протеинкиназа A управляет молекулярным переключателем, который управляет дифференцировкой псевдогифала дрожжей. Mol Cell Biol 22 (12): 3981–3993.
- 28. Галицкий Т., Салдана А.Дж., Styles CA, Lander ES, Fink GR (1999) Регулирование плоидности экспрессии генов.Наука 285 (5425): 251–254.
- 29. Verstrepen KJ, Jansen A, Lewitter F, Fink GR (2005) Внутригенные тандемные повторы генерируют функциональную изменчивость. Нат Жене 37 (9): 986–990.
- 30. Halme A, Bumgarner S, Styles C, Fink GR (2004) Генетическая и эпигенетическая регуляция семейства генов FLO порождает вариации клеточной поверхности у дрожжей. Ячейка 116 (3): 405–415.
- 31. Борнеман А.Р., Ли-Белл Дж. А., Ю Х., Бертоне П., Герштейн М. и др. (2006) Белки-концентраторы-мишени служат главными регуляторами развития дрожжей.Genes Dev 20 (4): 435–448.
- 32. van Dyk D, Pretorius IS, Bauer FF (2005) Mss11p является центральным элементом регуляторной сети, которая контролирует экспрессию FLO11 и инвазивный рост в saccharomyces cerevisiae. Генетика 169 (1): 91–106.
- 33. Gagiano M, van Dyk D, Bauer FF, Lambrechts MG, Pretorius IS (1999) Msn1p / Mss10p, Mss11p и Muc1p / Flo11p являются частью пути передачи сигнала ниже Mep2p, регулирующего инвазивный рост и дифференцировку псевдогифа в saccharomyces cerevisiaces.Mol Microbiol 31 (1): 103–116.
- 34. Lorenz MC, Heitman J (1998) Регуляторы псевдогифальной дифференцировки у saccharomyces cerevisiae, идентифицированные с помощью многокопийного супрессорного анализа в мутантных штаммах аммонийной пермеазы. Генетика 150 (4): 1443–1457.
- 35. Raj A, Peskin CS, Tranchina D, Vargas DY, Tyagi S (2006) Стохастический синтез мРНК в клетках млекопитающих. PLoS Biol 4: e309.
- 36. Belli G, Gari E, Piedrafita L, Aldea M, Herrero E (1998) Двойная система активатор / репрессор обеспечивает жесткую регулируемую тетрациклином экспрессию генов в почкующихся дрожжах.Nucleic Acids Res 26 (4): 942–947.
- 37. Каплан Н., Мур И.К., Фондуф-Миттендорф Ю., Госсетт А.Дж., Тилло Д. и др. (2008) ДНК-кодируемая нуклеосомная организация эукариотического генома. Nature 458 (7236): 362–366.
- 38. Conlan RS, Tzamarias D (2001) Sfl1 функционирует через корепрессор Ssn6-Tup1 и цАМФ-зависимую протеинкиназу Tpk2. Журнал молекулярной биологии 309 (5): 1007–1015.
- 39. Бергер С.Л., Пина Б., Сильверман Н., Маркус Г.А., Агапите Дж. И др.(1992) Генетическая изоляция ADA2: потенциальный адаптер транскрипции, необходимый для функционирования определенных доменов кислотной активации. Cell 70 (2): 251–265.
- 40. Song W, Carlson M (1998) Srb / белки-медиаторы функционально и физически взаимодействуют с репрессором транскрипции Sfl1. EMBO J 17 (19): 5757–5765.
- 41. Suka N, Suka Y, Carmen AA, Wu J, Grunstein M (2001) Высокоспецифичные антитела определяют использование сайта ацетилирования гистонов в дрожжевом гетерохроматине и эухроматине.Mol Cell 8 (2): 473–479.
- 42. Pelechano V, Garcia-Martinez J, Perez-Ortin JE (2006) Геномное исследование расстояний между ORF в saccharomyces cerevisiae. Дрожжи 23 (9): 689–699.
- 43. Lim HN, van Oudenaarden A (2007) Многоступенчатый эпигенетический переключатель обеспечивает стабильное наследование состояний метилирования ДНК. Нат Генет 39 (2): 269–275.
- 44. Де Лас Пенас А., Пан С.Дж., Кастано И., Олдер Дж., Крегг Р. и др. (2003) Связанные с вирулентностью поверхностные гликопротеины дрожжевого патогена Candida glabrata кодируются субтеломерными кластерами и подвергаются RAP1- и SIR-зависимому транскрипционному молчанию.Genes Dev 17 (18): 2245–2258.
- 45. Lorenz MC, Heitman J (1997) Рост псевдогифала дрожжей регулируется GPA2, альфа-гомологом G-белка. EMBO J 16 (23): 7008–7018.
- 46. Aparicio O, Geisberg JV, Struhl K (2004) Иммунопреципитация хроматина для определения ассоциации белков со специфическими геномными последовательностями in vivo. Curr Protoc Cell Biol Глава 17: Раздел 17.7.
- 47. Ян Б., Кирчмайер А.Л. (2006) Обход каталитической активности SIR2 для распространения белка SIR в saccharomyces cerevisiae.Mol Biol Cell 17 (12): 5287–5297.
- 48. Lam FH, Steger DJ, O’Shea EK (2008) Хроматин отделяет порог промотора от динамического диапазона. Nature 453 (7192): 246–250.
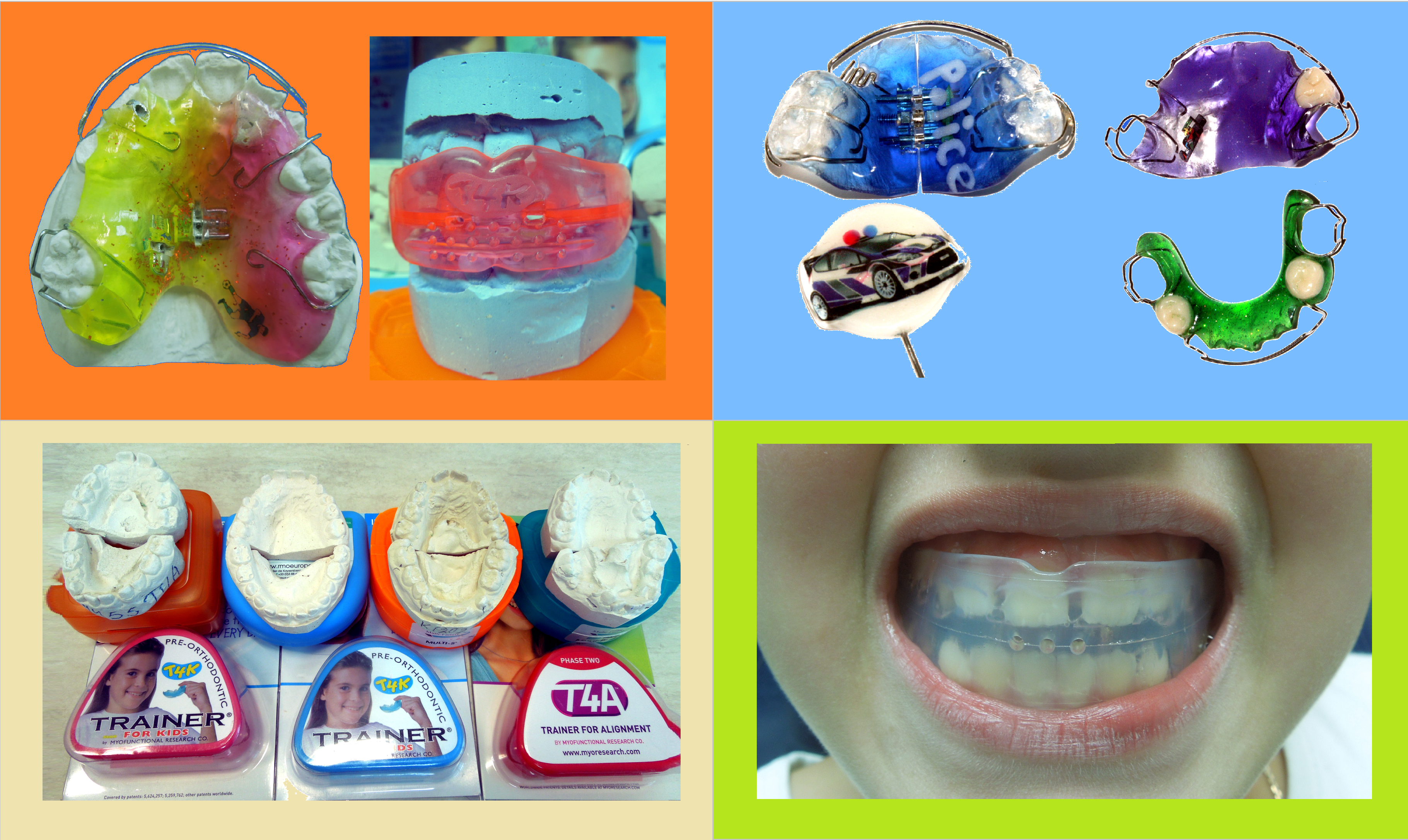 Curr Opin Microbiol 7 (4): 369–374.
Curr Opin Microbiol 7 (4): 369–374. Молекулярная ячейка 16 (4): 537–547.
Молекулярная ячейка 16 (4): 537–547.«Активность и природа активаторов плазминогена, связанных с этим случаем» Б. Исмаил, Л. Х. Чой и др.
Аннотация
В свежем молоке плазминоген, зимогенная форма плазмина (PL), является преобладающей формой. Следовательно, активаторы плазминогена (PA) могут вносить значительный вклад в активность PL в молоке.PA тканевого типа (tPA) и PA урокиназного типа (uPA) существуют в молоке; однако были получены противоречивые данные о том, какой тип PA наиболее тесно связан с мицеллами казеина. Мало что известно о факторах, которые могут привести к вариациям в индивидуальных действиях ПА. Таким образом, целью данной работы было изучить возможные факторы, которые могут повлиять на ассоциацию tPA и uPA с мицеллой казеина и их активность в дальнейшем. Активаторы плазминогена выделяли из образцов молока с различным количеством соматических клеток по 2 различным протоколам выделения.Определение активности uPA, tPA и PL проводили количественно после хромогенных анализов с использованием 2 различных субстратов и качественно с использованием специализированного додецилсульфата натрия-PAGE. Различные методы и условия выделения приводили к различиям в активности uPA, tPA и PL. Активность ПА урокиназного типа была значительно выше во фракциях ПА, выделенных из молока с высоким количеством соматических клеток, чем из молока с низким количеством соматических клеток. Результаты активности показали, что в пастеризованном молоке uPA может диссоциировать от соматических клеток и связываться с казеином.Более того, высокий уровень PL в изолированных фракциях PA способствовал значительному усилению активности PA. В целом, результаты подтвердили связь как uPA, так и tPA с мицеллой казеина; однако их количество, активность и молекулярная масса варьировались в зависимости от природы молока и методов разделения, при этом uPA является PA с большим потенциалом влиять на активацию плазминогена в молоке.
Рекомендуемое цитирование
Исмаил, Б., Л. Х. Чой, Л. М. Уэр и С. С.Нильсен. «Активность и природа активаторов плазминогена, связанных с мицеллой казеина». Journal of Dairy Science 89.9 (2006): 3285-3295.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
% PDF-1.5 % 1 0 объект > / OCGs [8 0 R] >> / Страницы 2 0 R / Тип / Каталог >> эндобдж 38 0 объект > / Шрифт >>> / Поля [] >> эндобдж 42 0 объект > поток 2021-06-05T15: 33: 36-07: 002006-09-30T11: 02: 46 + 08: 002021-06-05T15: 33: 36-07: 00uuid: 660aba65-f786-4421-a6ef-24752e7531a5uuid: ea8608d5- 1dd1-11b2-0a00-8800781dd8ffapplication / pdf конечный поток эндобдж 2 0 obj > эндобдж 33 0 объект > / Ресурсы> / Шрифт> / T1_1> / T1_2> / T1_3 46 0 R >> / ProcSet [/ PDF / Text / ImageB] / Properties> / XObject >>> / Type / Page >> эндобдж 28 0 объект > / Ресурсы> / Шрифт> / T1_1> / T1_2> / T1_3> / T1_4 46 0 R >> / ProcSet [/ PDF / Text / ImageB] / Properties> / XObject >>> / Type / Page >> эндобдж 18 0 объект > / Resources> / Font> / T1_1> / T1_2> / T1_3> / T1_4> / T1_5 46 0 R >> / ProcSet [/ PDF / Text / ImageB / ImageC] / Properties> / XObject >>> / Type / Page >> эндобдж 13 0 объект > / Ресурсы> / Шрифт> / T1_1> / T1_2 46 0 R >> / ProcSet [/ PDF / Text / ImageB] / Свойства> / XObject >>> / Type / Page >> эндобдж 5 0 obj > / Resources> / Font> / T1_1 46 0 R >> / ProcSet [/ PDF / Text / ImageB] / Properties> / XObject >>> / Type / Page >> эндобдж 43 0 объект > / ProcSet [/ PDF / Text / ImageC] / XObject >>> / Type / Page >> эндобдж 55 0 объект [61 0 R 62 0 R 63 0 R 64 0 R 65 0 R] эндобдж 56 0 объект > поток q 540.0594177 0 0 68.6011963 35.9702911 675.3988037 см / Im0 Do Q BT / T1_0 1 Тс 10 0 0 10 85,56995 558,99982 тм (1990; 50: 4957-4961.) Tj / T1_1 1 Тс -5.55699 0 Тд (Рак Res \ 240) Tj / T1_0 1 Тс 0 1 ТД (\ 240) Tj 0 1.00001 TD (Роберт Г. Ситрин, Маргарет Р. Гьетко, Керри Л. Коле и др.) Tj / T1_2 1 Тс 0 1 ТД (\ 240) Tj / T1_3 1 Тс 18 0 0 18 30 598,99994 тм (Линии глиомы) тиджей Т * (Активаторы и ингибиторы активаторов плазминогена человека) Tj Т * (Экспрессия гетерогенных профилей плазминогена) Tj ET 30 504 552 35 рэ 0 0 мес. S BT / T1_0 1 Тс 11 0 0 11 120.94202 511.99997 Тм (\ 240) Tj / T1_3 1 Тс -7,55696 1 тд (Обновленная версия) Tj ET BT / T1_2 1 Тс 10 0 0 10 141 503,99994 тм (\ 240) Tj / T1_0 1 Тс 23.17895 1 тд () Tj 0 0 1 рг -23.17895 0 Тд (http://cancerres.aacrjournals.org/content/50/16/4957)Tj 0 г 0 1.00001 TD (См. Самую последнюю версию этой статьи по адресу:) Tj ET BT / T1_2 1 Тс 10 0 0 10 30 483,99997 тм (\ 240) Tj 0 1 ТД (\ 240) Tj ET BT / T1_2 1 Тс 10 0 0 10 30 463,99997 тм (\ 240) Tj Т * (\ 240) Tj ET BT / T1_2 1 Тс 10 0 0 10 30 443,99997 тм (\ 240) Tj Т * (\ 240) Tj ET 30 329 552 115 рэ 0 0 мес. S BT / T1_0 1 Тс 11 0 0 11 120.94202 411,99997 тм (\ 240) Tj / T1_3 1 Тс -5.66901 1 тд (Оповещения по электронной почте) Tj ET BT / T1_0 1 Тс 10 0 0 10 295,4996 424 тм (относится к этой статье или журналу.) Tj 0 0 1 рг -15.44996 0 Тд (Подпишитесь, чтобы получать бесплатные уведомления по электронной почте) Tj ET BT 0 г / T1_0 1 Тс 11 0 0 11 120.94202 378.99994 тм (\ 240) Tj / T1_3 1 Тс -6.38997 1 тд (Подписки) Tj 0,556 1,00001 тд (Отпечатки и) Tj ET BT / T1_0 1 Тс 10 0 0 10 141 381,99994 тм (\ 240) Tj 13,46496 1 тд (.) Tj 0 0 1 рг -6.85098 0 Тд ([email protected]) Tj 0 г -6.61398 0 Тд (Отделение) Tj 0 1.00001 TD (Чтобы заказать перепечатку статьи или подписаться на журнал, свяжитесь с нами \ t Публикации AACR) Tj ET BT / T1_0 1 Тс 11 0 0 11 120.94202 356.99997 тм (\ 240) Tj / T1_3 1 Тс -5.66901 1 тд (Разрешения) Tj ET BT / T1_0 1 Тс 10 0 0 10 141 328.99988 тм (\ 240) Tj 0 1 ТД (Сайт с правами.) Tj 0 1.00001 TD (Нажмите «Запросить разрешения», чтобы перейти на страницу защиты авторских прав \ Центр Рэнсиса \ (CCC \)) Tj 23.17895 1 тд (.) Tj 0 0 1 рг -23.17895 0 Тд (http://cancerres.aacrjournals.org/content/50/16/4957)Tj 0 г 0 1 ТД (Чтобы запросить разрешение на повторное использование всей или части этой статьи, используйте это li \ nk) Tj ET BT / T1_0 1 Тс 9 0 0 9 258.\ q
ПРАЙМ PubMed | Экспрессия гетерогенных профилей активаторов плазминогена и ингибиторов активаторов плазминогена линиями глиомы человека
Citation
Sitrin, R G., et al. «Экспрессия гетерогенных профилей активаторов плазминогена и ингибиторов активаторов плазминогена линиями глиомы человека». Cancer Research, vol. 50, нет. 16, 1990, стр. 4957-61.
Ситрин Р.Г., Гетко М.Р., Коле К.Л. и др. Экспрессия гетерогенных профилей активаторов плазминогена и ингибиторов активаторов плазминогена линиями глиомы человека. Cancer Res . 1990; 50 (16): 4957-61.
Ситрин, Р. Г., Гьетко, М. Р., Коле, К. Л., МакКивер, П., и Варани, Дж. (1990). Экспрессия гетерогенных профилей активаторов плазминогена и ингибиторов активаторов плазминогена линиями глиомы человека. Cancer Research , 50 (16), 4957-61.
Sitrin RG, et al. Экспрессия гетерогенных профилей активаторов плазминогена и ингибиторов активаторов плазминогена линиями глиомы человека. Cancer Res., 15 августа 1990; 50 (16): 4957-61. PubMed PMID: 2379161.
TY — JOUR T1 — Экспрессия гетерогенных профилей активаторов плазминогена и ингибиторов активаторов плазминогена линиями глиомы человека. AU — Ситрин, R G, AU — Гетко, М. Р., AU — Коле, К Л, AU — Маккивер, П, AU — Варани, Дж., PY — 1990/8/15 / pubmed PY — 1990/8/15 / medline PY — 1990/8/15 / entrez СП — 4957 EP — 61 JF — Исследования рака JO — Cancer Res ВЛ — 50 ИС — 16 N2 — Экспрессия активности фермента активатора плазминогена (PA) считается одним из механизмов, с помощью которого злокачественные клетки вызывают перицеллюлярный протеолиз стромальных структур во время имплантации и тканевой инвазии.В этом исследовании изучались четыре клеточные линии, полученные из глиом человека, чтобы установить, какие ферменты PA и ингибиторы PA определяют уровень секретируемой активности PA. Был использован плазминоген-зависимый эстеролитический анализ, и было обнаружено, что две линии (U251 и U373) секретируют высокие уровни активности PA, в то время как активность PA не определялась в кондиционированной среде от оставшихся двух линий (U138 и LM). PA, продуцируемый U251 и U373, разделился на отдельные полосы, соответствующие высокомолекулярной урокиназе (Mr 54000) при зимографии казеин-плазминоген.Нозерн-блоттинг продемонстрировал высокие уровни мРНК PA урокиназного типа (uPA) в U251 и U373, а также значительно более низкий уровень сообщения uPA в LM. U251 и U373 также содержали мРНК для PA тканевого типа (tPA), хотя активность секретируемого tPA не была продемонстрирована зимографией. Линия U138 содержала по существу неопределяемые уровни мРНК для uPA или tPA. U138 также обладал уникальной секретирующей активностью ингибитора PA и содержал высокие уровни мРНК для ингибитора 2 PA, которые не наблюдались ни в одной другой линии.Все клеточные линии содержали мРНК ингибитора 1 PA, с существенно большей экспрессией в линиях U138 и LM, чем в U251 и U373. Ни одна из линий не секретировала измеряемой антиплазминовой активности. Мы пришли к выводу, что среди клеток глиомы человека существует значительная гетерогенность в экспрессии ферментов PA и ингибиторов PA. Скоординированная регуляция этих белков, вероятно, определяет активность секретируемого PA и, как следствие, роль активации плазминогена в имплантации и инвазии опухоли. SN — 0008-5472 UR — https: // neuro.unboundmedicine.com/medline/citation/2379161/expression_of_heterogen_profiles_of_plasminogen_activators_and_plasminogen_activator_inhibitors_by_human_glioma_lines_ L2 — http://cancerres.aacrjournals.org/cgi/pmidlookup?view=long&pmid=2379161 БД — ПРЕМЬЕР DP — Unbound Medicine ER —
Активаторы транскрипции выводят порядок из хаоса
Поскольку транскрипционные факторы AD имеют тенденцию быть структурно неупорядоченными и часто способны взаимодействовать со многими неродственными коактиваторами, биохимики давно задаются вопросом, как они достигают специфичности; то есть, как факторы транскрипции так хороши для привлечения нужного коактиватора в нужное место в нужное время? Лаборатория Хана в отделе фундаментальных наук пытается ответить на этот вопрос, изучая, как фактор транскрипции Gcn4, датчик статуса питательных веществ, взаимодействует с Med15, компонентом большого комплекса коактиваторов, известного как медиатор.
В предыдущей работе лаборатория Хана обнаружила, что интерфейс Gcn4-Med15 на удивление динамичен или «нечеткий». «Нечеткие взаимодействия — это взаимодействия, в которых партнеры по связыванию не принимают фиксированную конформацию, а вместо этого образуют множество различных конформаций», — объясняет доктор Лиза Таттл, научный сотрудник лаборатории Хана, которая также является членом лаборатории Клевит в Отделение биохимии UW.
Первоначальное исследование Gcn4-Med15 было сосредоточено на взаимодействии между одним Gcn4 AD и одним активатор-связывающим доменом Med15 (ABD).Чтобы определить, является ли нечеткое связывание особенностью комплекса Gcn4-Med15 в целом, доктор Таттл и ее коллеги из лабораторий Хана и Клевита решили охарактеризовать взаимодействия между обоими AD Gcn4 и всеми четырьмя ABD Med15. Их результаты, недавно опубликованные в Cell Reports , , , показывают, что Gcn4 и Med15 действительно достигают сильного и функционального взаимодействия посредством множества слабых, динамических контактов AD-ABD.
Во-первых, исследователи использовали изотермическую калориметрию (ITC) и поляризацию флуоресценции (FP) для измерения силы связывания между фрагментами Gcn4 и Med15, содержащими 1-2 AD и 1-4 ABD, соответственно.Они обнаружили, что все парные взаимодействия между отдельными AD и отдельными ABD были слабыми, но взаимодействия постепенно усиливались по мере добавления большего количества AD или ABD.
Основываясь на своих данных, авторы представили два возможных сценария для интерфейса между полноразмерными Gcn4 и Med15: либо он может оставаться нечетким при добавлении компонентов, либо комплекс может в конечном итоге превратиться в жесткую структуру. Чтобы различать эти возможности, авторы химически сшили Gcn4, содержащий оба AD, с Med15, содержащим все ABD, и проанализировали связи с помощью масс-спектрометрии.Они наблюдали связи почти между каждой парой AD-ABD, даже между теми, чье сродство было ранее показано как низкое, предполагая, что события слабого связывания не конкурируют с другими взаимодействиями в контексте полноразмерных белков. Этот результат согласуется с тем, что большинство или все взаимодействия AD-ABD являются временными.
В то время как эксперименты по химическому сшиванию продемонстрировали, что взаимодействия AD-ABD имеют тенденцию быть динамическими, они напрямую не касались того, принимают ли индивидуальные интерфейсы AD-ABD уникальные или нечеткие конформации во время каждого события связывания.Чтобы ответить на этот вопрос, д-р Таттл и ее коллеги использовали ядерный магнитный резонанс (ЯМР) для сравнения свойств комплексов AD-ABD изолированно и внутри всего комплекса. Помечая AD тяжелыми изотопами азота и углерода, они могли определить, стал ли его полипептидный каркас, который по своей природе неупорядочен, более упорядоченным при добавлении одного или нескольких ABD. Как и ожидалось, нарушенные области скелета AD соответствуют гидрофобным кластерам, ранее участвовавшим в связывании ABD; никаких новых взаимодействий не наблюдалось в комплексах, содержащих все ABD, которые не присутствовали в смесях AD с отдельными ABD.
Несколько неожиданно, одни и те же области AD были нарушены независимо от того, какой ABD был добавлен, что позволяет предположить, что AD связываются независимо от идентичности ABD. В соответствии с этой идеей авторы продемонстрировали в последующих экспериментах ЯМР, что оба AD вызывают одинаковые изменения в ABD1 и ABD2 и конкурируют друг с другом за сайты связывания ABD1 и ABD2. Этот результат был неожиданным, поскольку было обнаружено, что ABD1 и ABD2 имеют очень разные карманы связывания AD.
Чтобы изучить ориентацию (ориентации), с которой AD связывают ABD, исследователи прикрепили парамагнитные зонды к каждой AD в разных местах и провели эксперименты со спиновой меткой ЯМР.Если AD всегда принимает одну и ту же конформацию в данном комплексе AD-ABD, авторы ожидают увидеть разницу в сигнале от зондов, размещенных на противоположных концах AD. Однако они наблюдали аналогичное поведение зонда независимо от его положения, что указывает на то, что AD не выказывали предпочтения определенной ориентации.
В целом, подробные исследования комплекса Gcn4-Med15 лабораториями Hahn и Klevit показывают, что многие слабые взаимодействия действительно могут синергизировать для достижения связывания с высокой аффинностью без принятия определенного интерфейса.Этот результат подчеркивает представление о том, что слабые белок-белковые взаимодействия могут вносить важный вклад в биологические процессы. Поскольку факторы транскрипции часто взаимодействуют с несколькими коактиваторами, а коактиваторы имеют тенденцию иметь несколько ABD, нечеткие интерфейсы связывания могут обеспечить большую гибкость в использовании различных комбинаций AD-ABD, облегчая тем самым сложные механизмы регуляции генов.
Таттл Л.М., Пачеко Д., Варфилд Л., Ло Дж., Раниш Дж., Хан С. и Клевит РЭ.Специфичность Gcn4-медиатора опосредована большим и динамичным нечетким белково-белковым комплексом. Cell Reports. 20 марта 2018 г.
Фред Хатч / члены Консорциума по раку из UW Стив Хан (Фред Хатч) и Рэйчел Клевит (UW) внесли свой вклад в эту работу.